光合成の生理生態学講座
はじめに(200101 29)
光合成能力の定義(200101 29)
光合成能力の種間差のパタン(200101 29)
C3とC4(200102 06)
ギャップ依存種と林床種(200102 06)
栄養条件(200104 21)
葉の特性(200104 21・2004 0911追加)
標高(200104 21)
光合成能力と窒素含量の関係の種間差(200105 12)
種間差をもたらす候補(200105 20・2004 0911追加)
種間差を調べる (200106 16・200106 28・2004 0911追加)
種間差の生態学的意義(200106 28・2004 0911追加)
異なる代謝経路を持つC3とC4ならともかく、同じC3植物なら、同じ光合成能力を持っていそうです。しかし実際のところは、同じ条件で育てたC3植物種でも10倍以上違う光合成能力を持っていたりします。光合成能力が高い、というのはなんとなく都合がいいような気がしますが、光合成能力が低い植物は生産効率が悪そうで、とっくの昔に淘汰されていいような気がします。何が良くて光合成能力の低い植物が生きていけるのでしょうか? この項では、1)どういう植物で光合成能力が低いのか、2)光合成能力の違いをもたらす生理学的原因は何か、3)光合成能力が低いことの適応的意義は何か、について、わかっているところまで書きたいと思います。
その前に、「光合成能力(photosynthetic capacity)」は何かを定義しなければいけません。しかし、論文によって定義はまちまちで、注意が必要です。
まず、どのような条件で光合成速度を測定するかがポイントです。最も一般的に使われているのは、「生育条件と同じ大気組成(酸素および二酸化炭素濃度のこと)・生育条件に近い温度・充分に高い湿度(一般に、VPD<1kPa)・飽和光強度で測定した光合成速度」です。本項でもこの定義にしたがいます。別の定義として、最適(に近い温度)で測定したものとか、25度で測定したものを使う場合もあります。前者は理想的ですが、いちいち光合成の最適温度を探すのはかなり面倒なので、あまり使われません。後者はかなり多く使われます。これは、慣習的にこの温度での測定例が多く、他の研究との比較が容易なためです。他に、飽和二酸化炭素濃度で測定した光合成速度を光合成能力と定義する場合もあります。これは生育二酸化炭素濃度が異なる場合によく使われます。
次に、光合成能力を何当たりで表すかがポイントです。最も多く使われているのは葉面積あたりですが、近頃増えてきたのが乾燥重量あたりで表したものです。近頃増えてきた理由はPeter Reichのグループが乾燥重量当たりの光合成能力を頻繁に使っているからです。私のように葉面積あたりの光合成能力を使い慣れている人間にはなかなか理解しがたい使い方ですが、ともかくReichらの影響は大きいので無視できません。このため、この項でも両方を使います。以下、葉面積あたりの光合成能力はAarea、乾重あたりの光合成能力をAmassとします。なかなか混乱したものになると思いますが、よろしくおつきあい下さい。
ついでに。「窒素当たりの光合成速度」を光合成能力とする著者もまれにいますが、本項ではこれを「光合成の窒素利用効率(photosynthetic nitrogen use efficiency, 以下PNUE)」と呼び、光合成能力とは区別します。ただ、PNUEも本項では重要な性質として扱います。それは、PNUEは、同一種内ならば生育環境が変わってもそれほど大きくは変化せず、種固有の特性の一つとして考えることができるためです。Aareaが生育条件によって変化するのはこちらで説明した通りです。
光合成能力の種間差のうち、古くから知られていたのは「木本より草本でAareaが高い」というものです。Larcher の教科書にも、たぶん初版から記述されていたと思います(私が持っているのは2版。最近日本語に訳されたのは第5版)。例えば、草本ならばC3であっても30-40μmol m-2 s-1に達することは普通に見られます。一方、木本では、20μmol m-2 s-1を超えることはまれで、ほとんどは10μmol m-2 s-1前後でしょう。
この違いが生理生態学における大きなテーマの一つとなったのは、Field and Mooney (1986) のレビューです。Mooney のグループは、70年代から光合成と窒素の相関に着目していました。同一種内ではAareaと葉面積あたりの窒素含量(Narea)の間に高い相関があることがわかっていました。Field and Mooney (1986) では、それまでの光合成と窒素の関係についての文献を集め、10数種のデータを一つのグラフにプロットしてみました。すると、種が異なるとこの関係は大きく異なることに気がつきました。木本種と草本種では、同じNareaで比較しても草本のほうが高いAareaを持ちます。さらに、彼らは光合成と窒素の両方を乾燥重量あたりで表してみました(Amass, Nmass)。すると、葉面積あたりではばらばらだったグラフが、ほぼ一本の線にのりました。こういった傾向がなぜ現れるのか?が以降の研究者たちの興味になりました。
これまで、様々な研究者が生態学的特性(しばしばfunctional groupと呼ばれる)と光合成能力・PNUEの間の関係を調べてきました。では、どのような傾向があるのか、これまで報告された研究例を私が知る限りご紹介しましょう。
C4植物は一般に高い光合成能力を持っていることがしられています。同一条件で育てれば、C3植物よりもC4植物で光合成能力が高くなります。また、PNUEもC4で高くなります。PNUEの違いは古くから興味が持たれており、いくつか研究があります(Brown 1978, Sage and Pearcy 1987, Anten et al. 1995a)。
ただし、全てのC4植物が高い光合成能力を持つわけではないという話も耳にします。C4には3つのサブタイプがあり、NADP-ME型、NAD-ME型、PCK型と呼ばれています(中間代謝産物が若干異なる)。数年前に、タイプによって光合成能力が違うという話を(本当に話だけ)聞いたことがあります。文献は知らないので、本当かどうかは定かではありません。
CB Field のグループは80年代の中頃にメキシコの熱帯雨林でコショウ属の様々な生態的特性を持った種について光合成特性などを比較しています(Walters and Field 1987, Field 1987など)。その一連の仕事の中に、6種の光合成能力を比較した、というものがあります(Chazdon and Field 1987)。その研究では、明るいところを好んで生える植物のほうがAmass、Aareaともに高い値を示しています。ただし、この研究では現地で光合成速度を測定していますので、Aareaの違いは生育環境の違いを反映している可能性が高いです(実際、この論文のテーマも光環境と光合成特性の関係です)。ただ、種内では生育環境が変わってもあまり変化しないと考えられているPNUEは明るいところを好んで生える種のほうが高い値を持っているようです。
ヨーロッパではEllenberg (1979) という植物社会学系の論文で、ニッチに基づいた植物の分類がされているのだそうです(残念ながらドイツ語のため読んでいません)。例えば、栄養条件なら、最も富栄養な条件を好む種は10、最も貧栄養な条件を好む種は1というように10段階で評価されます(N numberといいます)。栄養条件以外の環境などでも評価されており、ある種がどういう条件を好むのかが一目瞭然になっているようです。
H. Poorter のグループは様々な草本24種を用い、成長解析、光合成、呼吸、構成コスト、水分生理など様々なテーマについて徹底した種間比較を行いました。彼らの材料ではN numberが高い種ほどRGRが高いという結果が出ています(Poorter and Remkes 1990)。彼らの研究のうち、Poorter et al. (1990) が炭素・窒素経済について解析しており、光合成のデータも載っています。AareaとRGRの間に相関はありませんでしたが、AmassとPNUEはRGRと相関があり、富栄養条件を好む植物ほどAmassとPNUEが高いと言えそうです。
種によって葉の特性が大きく違います。光合成能力と相関があることが古くから知られていた葉の特性は、葉の寿命でしょう。
葉の寿命が短い種ほどAmass, Aarea, PNUEとも高いことが知られています。非常に多くの研究例があり、よく知られたレビューはChabot and Hicks (1982) です。近年ではReichらのグループのReich et al. (1991) 以降のシリーズが代表的な研究だと思います(葉の寿命についてはまた別に書きたいと考えています)。
Reich et al. (1991) は(たぶん)多くの研究者に影響を与えた研究です(少なくとも私はたいへん大きな影響を受けました)。彼らが行ったのは、アマゾンの熱帯雨林のあちこちで異なる種の葉の光合成・葉面積あたりの葉重(LMA)・葉寿命・窒素濃度を測定する、というものです。この論文で彼らが示したのは、これらの間に非常にきれいな(美しいと表現して良いくらいの)相関があったことです。葉の寿命が長い種ほどAmass、PNUEが低く、LMAが高く、乾燥重量あたりの窒素濃度(以下Nmass)が低いということがクリアに示されました。ちなみに、Areaや葉面積あたりの窒素含量(以下Narea)はあまり違いがなかったようです。
Reichらはこの関係がどこまで共通なのかを探ることにしました。熱帯雨林の植物(Reich et al. 1994, Reich and Walters 1994, Ellsworth and Reich 1996)のほかに、温帯に生育する落葉樹と常緑針葉樹の比較(Reich et al. 1995)、そして様々な気候帯(彼らはbiomeという言葉を使っています)での測定を比較することにより、1991年のアマゾンで見られたような関係、すなわち、葉の寿命が長い種ほどAmass、PNUEが低く、LMAが高く、乾燥重量あたりの窒素濃度(以下Nmass)が低いということを見いだしました(Reich et al. 1997, 1998a, 1999)。この傾向は、それぞれのパラメータを対数表示すると一本の線に見える、というものです。よくよく見てみるとバイオーム間などでは有意に関係が違う(回帰直線の切片などが異なる)ようですが、まあ、傾向としては同一線上にある、ということです。ついでに彼らは葉の呼吸速度も光合成速度と同様な傾向があることを示しています(Reich et al. 1998b)。
この葉の形質の傾向探しは行き着くところまで行き着き、2500種で相関関係を探る、という研究が行われました。I Wright, PB Reich, M Westobyが中心になり、Glopnet(Global plant trait network)が組織され、これまでの既・未発表データをかき集め、解析が行われました(Wright et al. 2004)。2500種というのは維管束植物27万種の約1%ということになります。また、地球上様々な気候・地点からデータをとる工夫がされて、南極大陸を除く全ての大陸、地球上で陸上植物が生育するほとんどの平均気温-降水量の組み合わせを含んでいます。私のデータも25種ほど使われていて(清澄とキナバル)、この研究に対する私の貢献は1%ということになります。ただ、2500種といっても、光合成・呼吸・窒素・リン・葉寿命がそろったデータというのはそう多くはありません。私のデータも呼吸とリンは入っていませんし。この2500種で行われた解析でも、基本的にはReichらと同様の結果が得られています。主成分分析を行うと、葉の特性は多次元空間の中の一本の回帰直線上で75%のばらつきを説明することができました。この一本の線の端にはAmass、Rmass(乾重あたりの呼吸速度)、Nmass、Pmassが高く、LMAが低く、葉寿命が短い種が集まり、もう一端には逆の性質の種が集まりました。つまり、種によって葉の性質は大きく異なるけれど、それはばらばらに違うわけではなく、一本の線上に集まるのだということがわかったわけです。
一本の線にのる、とは言いましたが、この関係は気候によって若干変わるようです。Wright and Westoby (2002) は葉の寿命とLMAの関係が、オーストラリアにおいて土地の栄養条件には依存しないが降水量によって変わることを示しました。Glopnetの結果もこれを支持しています。同じLMAで比べると、降水量が低い場所ほど葉の寿命が短くなります。これを説明する理屈は私にはよくわかりません(Wrightは気孔コンダクタンスなどを使って説明を試みているが私には納得行かない)。ただし、この違いも全体から見ると大きくはなく、おおざっぱには「一本の線上にのる」ということは充分言えるようです。
20年くらい前までは野外で光合成速度を測定するというのは絶望的に難しいことでした。スタンフォード大のグループなんかは車一台にどでかい測定機器をのっけて測定をしていたようですが、まあ誰にできるものではなかったことは確かです。ましてや高山に生えている植物の光合成なんて・・・。ところが80年代に入ってからは比較的軽量の携帯式光合成測定装置が登場し、誰でも野外で光合成速度を測定できるようになります。
標高間比較についてはこれまで様々な研究が行われていますが、必ずしも多くはありません。Korner and Diemer (1987) は近縁の植物種を選んで10くらいのペアを作り、現地での光合成速度を比較しました。傾向として言うと、標高が高いほどPNUEやAmassが低いものの、NareaがPNUEの低さを補償してAareaは変わらない、という結果を出しています。Friend et al. (1989) は広い標高範囲に生育するツツジ科の2種を調べ、PNUEが高標高で低いことは示していましたが、Amass、Aareaは・・・どうだったかな?(手元に論文がないのでよくわからない) Vitousek et al. (1990) はMetrosideros polymorphaという植物のPNUEが高標高で低いことを示しています。ただ、このMetrosideros polymorphaはよく使われる材料で、標高が変わってもPNUEやAareaは変わらないよ、というデータもある(Meinzer et al. 1992)のでよくわかりません。Terashima et al. (1993) はヒマラヤで測定を行い、4300mに生育している植物のAareaやPNUEは低地の植物とそれほど変わらないことを報告しています。
最近我々もマレーシア・キナバル山(標高4095m)で測定を行いました(Hikosaka e al. in press)。我々の視点はこれまでの研究とちょっと違って、特定の種に着目するのではなく、その標高の植生を代表する種に着目しました。PNUEやAmassは明らかに高標高の植物で低く、AareaもAmassほどではありませんが高標高で低い、という傾向が見られました。
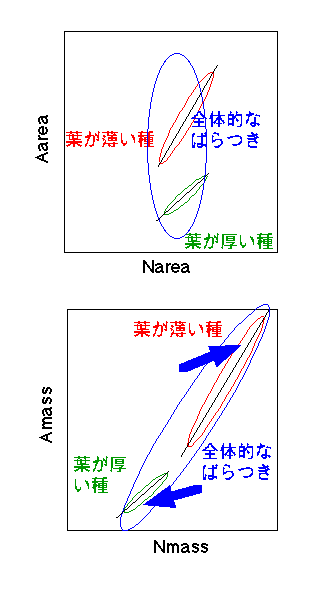
さて、この「光合成の生理生態学講座」では、あちこちで光合成能力と窒素含量の間に高い相関があることを書いてきました。この高い相関はあくまで同一種内のもので、種間では異なります。種間で窒素と光合成の関係を比べると、どうなるでしょうか? Field and Mooney (1986) は文献サーベイを行った結果、AmassとNmassの間には種に関わらず強い相関があるが、AareaとNareaの間の関係は種間で違う、と述べています。なぜ「乾重あたり」と「葉面積あたり」で話が違うのでしょうか?
まず、思い出していただきたいのは、Reichらが示した一般則、「Amass、Aareaが低い植物は、葉の寿命が長く、LMAが高く、Nmassが低い」というものです。これと、「種内では、乾重あたりでも葉面責あたりでも窒素含量と葉面積の間に相関がある」ということを合わせて考えてみましょう。まず、AreaとNareaの関係を見てみましょう。私がこれまで見てきた限りでは、おおざっぱにいって、生育条件が同じならばNareaの種間差は小さいようです(全く同じなわけではない)。しかし、同じ窒素含量ならば、LMAが低い種(図中では「葉が薄い種」)が高いAareaを持ちます。結果として、全体的なばらつきは縦長になります。とても相関があるとは言えません。これをふまえて、AmassとNmassの関係を見てみましょう。Amassとは、AreaをLMAで割ったものです(Amass=Aarea/LMA)。ついでに、Nmass=Narea/LMAです。ここで、Areaが高い種はLMAが低い、という傾向があることを思い出して下さい。ってことは、Aareaが高い種では、分母(LMA)が小さいのでAmassやNmassが相対的に高く、Aareaが低い種ではAmassやNmassが低くなります。そうすると、「葉面積あたり」のときは縦長だったばらつきが、右肩上がりになり、きれいな相関になる、ということです(下図)。
ここで注意していただきたいのは、Amass-Nmassの相関がきれいだということがPNUE(窒素あたりの光合成、=Amass/Nmass)一定を意味しない、ということです。Amass-Nmassの関係は直線的ですが、X切片がプラスの値になります。これは、Nmassが低い種ほどPNUEが低いことを意味します。
FieldらやReichらが示したAmass-Nmassの関係は本当にきれいです。Reichらのデータでは100を超える種がほぼ一本の線にのっています(対数グラフなので、ばらつきが見えにくくなっているからでもありますが)。しかし、なぜこのような関係があるのか、そして、この関係にどんな意味があるのかはまだ推測の域を出ていません。以下では、ここまでわかっていることを書くことにしましょう。
AareaやAmassは種内でも生育条件によって大きく変化します。しかし、この変化はNareaやNmassの違いで説明できることが多いです。つまり、多くの場合種内では光合成能力と窒素含量の関係は一本の「線」で表すことができ、生育条件によって線のどこにのるかが変わってきます。種間差の場合は、この「線」が変化することになります。そこで、生理学的なメカニズムとして種間差を考えるときは、AareaよりはPNUEやAarea-Nareaの回帰直線を比べるほうがわかりやすくなります。ここからは、PNUEの違いを見ていくことにしましょう。
まず、なぜ種内では光合成能力と窒素含量に高い相関があるのかを思い出して下さい。葉の窒素の大半は光合成系の窒素です。すなわち、葉の窒素量が多いことは光合成系のタンパク質量が多いことを意味し、高い光合成能力を実現できるわけです。では、同じ窒素量を持っていても光合成能力が違うのはなぜでしょうか?
光合成でCO2を固定する反応は酵素によって触媒される反応です。したがって、酵素の反応速度がどのように異なるかを考えれば済む話です。反応速度は、イ)基質の量、ロ)酵素の量、ハ)酵素のキネティクス(VmaxやKm)で決まります(これ以外の要因は、全て上の3つのどれかを介した間接的な効果です)。最初の反応を触媒する酵素はRuBPCaseですから、RuBPCaseについて考えます。RuBPCaseの基質はRuBP、CO2、O2の3つですが、ここでは光飽和の状態を考えますので、RuBPは飽和していると見なせます。また、O2の変化も考えなくていいでしょう。ということで、基質はCO2のみを考えます。
Field and Mooney (1986) は、PNUEの種間差の原因は以下の4つに分けられることを指摘しました。1)葉緑体内までのCO2拡散、2)葉の窒素のどれだけを光合成系に分配するか、3)光合成系の窒素をどのように分配するか、4)RuBPCaseの比活性(酵素の量あたりの反応速度)、です。このうち、1はイに、2と3がロ、4がハに相当します。
1の「葉緑体内までのCO2拡散」は、要するに、葉緑体までCO2が入っていきにくくなると葉緑体内のCO2濃度が下がり、光合成速度が下がる、というものです(CO2と光合成の関係はこちら。CO2の拡散についてはこちら)。気孔と細胞壁が主な律速段階と考えられています。
2と3は、葉の窒素量とRuBPCase量の関係を意味します。同じ窒素量を持っていても、RuBPCaseが少なければ、光合成能力が下がってしまいます。2と3の違いは、前者が葉全体の窒素の分配で、後者が光合成系内の窒素の分配ということです。後者は、例えば集光系に多く窒素を分配した結果光合成能力が下がることを想定しています(葉内の窒素分配についてはこちらとこちら)。
4は、RuBPCaseでいうと、VcmaxやKcなどの酵素活性パラメータの種間差を考えています。
ちなみに、Poorter and Evans (1998) はFieldらに加えて以下の要因を挙げています:5)光吸収率、6)Day respiration、7)光飽和点の違い。5と6は、Poorterらは生育環境におけるPNUE(必ずしも光飽和ではない)を考えているために挙げています。このページでは飽和光下のPNUEを考えるので無視します。6は、みかけの光合成速度は総光合成速度から呼吸速度を引いたものである、と考えると影響する可能性があることになります。Day respirationについてはこちら。
なお、Reich et al. (1997) では「葉が厚い場合、光が奥まで届かないため、葉の裏側付近の葉緑体が光律速になり、光合成能力が低いかもしれない」という考察をしています。ちなみに根拠として引用しているのがTerashima and Hikosaka (1995) なのですが、我々はそんなことは書いていませんし、そういうことが(全くないとはいわないけど)種間差をもたらす大きな要因だとは考えていません・・・というはずだったのですが、Green and Kruger (2001) が驚愕の結果を発表していました。彼らは落葉広葉樹と常緑針葉樹の光合成速度を測定しました。測定チャンバーを工夫することにより、光を上からも下からも当てられるようにし、光合成速度を測定しました。上から1800μmol m-2 s-1の光をあてたときに比べ、上下両方から1800μmol m-2 s-1の光(合わせて3600)をあてたときにどれだけ光合成速度が促進されたかを比較すると、落葉広葉樹では0-20%程度しか光合成速度が促進されなかったのに、常緑針葉樹では、多い葉では50%もの促進が見られました。促進の大きさはLMAときれいな正の相関がありました。このことは、常緑針葉樹では「葉が厚い場合、光が奥まで届かないため、葉の裏側付近の葉緑体が光律速になり、光合成能力が低い」ということを示唆しています。今のところこのような報告はGreen and Kruger (2001) にしかありませんので、どれだけ普遍的なのかはわかりません。しかしこの可能性を安易に排除してはいけないようです。
PNUEに種間差があることを最初に大きく指摘したのはField and Mooney (1986) のレビューです。次に、Evans (1989) のレビューがその原因について、より生化学的な知見に基づいた考察を行いました。しかし、Evansのレビューはいろいろな論文のデータを拾ってきての比較ですし、それまでに行われていた研究のほとんどがPNUEの高い農作物ばかりを対象としているので、それほど意味あるものではありませんでした。
とはいえ農作物では東北大の牧野先生が重要な仕事をしています(Makino et al. 1988)。この仕事ではイネとコムギのPNUEを比較しているのですが、両者はほぼ等しいPNUEを持ちます。しかし、イネは気孔コンダクタンスが高く、窒素あたりのRuBPCase量が高いのに対し、コムギはRuBCaseの比活性が高いことがわかりました。つまり、PNUEが等しいからといって上に挙げたそれぞれの要因が等しいとは限らない、ということです。
一つの実験の中でPNUEが違う原因を最初に調べたのはLloyd et al. (1992) です。この研究の作業仮説は「葉内細胞間隙から葉緑体までのCO2拡散抵抗の違いがPNUEの種間差をもたらす」でした。Lloydは当時キャンベラにいましたが(今も?)、キャンベラには葉内のCO2拡散を測定するシステムがあり(こちらを参照)、このシステムを使って種間差の問題を考えようとしたのです。Lloyd et al. (1992) が使ったのは、PNUEが高いモモ(落葉樹)とレモン、マカダミアンナッツ(常緑樹)です(これらもまあ農作物なんですが)。彼らはマカダミアンナッツで葉緑体CO2濃度が低いことを確かめましたが、それだけではPNUEの違いは説明できないことを示しました。さらに、ガス交換特性からRuBPCase量を推定し、窒素あたりのRuBPCase量の違いのほうが大きいことを示唆しました。
私は草本と常緑広葉樹の比較、ということで、シロザとシラカシの比較をしました(Hikosaka et al. 1998)。シラカシはシロザの半分のPNUEしかもちません。RuBPCase量・比活性の測定、ガス交換特性、13C含量の測定から、RuBPCase量の違いが約20%、葉内細胞間隙CO2濃度の違いが約10%、比活性の違いが約10%、葉緑体内のCO2濃度の違いが約5%で合計約50%の違いをもたらしていると結論しました。つまり、ある一つのパラメータの違いがそのままPNUEの違いになっているのではなく、いろんなパラメータの小さな違いの積み重ねがPNUEの違いを大きくしているのだと考えました。
で、Poorter and Evans (1998) です。Poorterは「草本24種の様々な性質の種間差」を徹底的に調べた研究者としてよく知られています。この論文もある意味そのシリーズの一つかもしれませんが、使った材料はシリーズで使ってきた材料とは違い、木本も含んでいます。種数も10種と少なくなっています。「異なるSLA(LMAの逆数)の葉を持つ種のPNUEの比較」というのが彼らの論文のタイトルです。彼らは10種の植物を2つの光条件で比較し、それぞれの条件においてのPNUEの違いの原因を調べています。彼らが測定したのは、RuBPCase量・chl量・ガス交換特性です。その結果、弱光条件では光合成系タンパク質への窒素分配の違いが、強光条件ではRuBPCaseあたりの光合成速度の違いが最も大きな原因であった、としています。ここでいうRuBPCaseあたりの光合成速度の違いは、葉緑体CO2濃度の違いと比活性の違いの二つの可能性がありますが、彼らは比活性は測定していないのでわかりません。なお、「最も大きな原因であった」と書きましたが、どちらもPNUEの違いの半分しか説明しておらず、他にも光合成系内の窒素分配なども少しづつ影響しているようです。
Westbeek et al. (1999) は異なる標高にハビタットをもつPoa属の4種を実験室で育ててPNUEを比較しています。彼らの測定項目もPoorter and Evans (1998) とほぼ同じです。高標高に生えている種ほどPNUEが低いのですが、この研究では光合成系に分配される窒素の割合が一番重要だ、としています。
葉内CO2濃度の違いにはおそらく多くの研究があるのではないかと思います。様々な植物の比較をした研究として知られているのはYoshie (1986) とPoorter and Farquhar (1994) です。前者は温帯草原・落葉樹林の様々な種の光合成速度と気孔コンダクタンスを比較し、両者が正比例している、つまり葉内CO2濃度はほぼ一定であることを示しています。Poorter and Farquhar は例の「24種の比較」のシリーズで気孔コンダクタンスについて調べており、やはり葉内CO2濃度にはこれといった種間差がないことを示しています。
RuBPCaseの比活性の比較の研究は、農作物ではかなりあるようですが、野生植物では多くないのが現状です(Evans 1989を参照)。必ずしもPNUEが低い植物で比活性が低いとは限らない、と言っている論文もあります(Vu and Yelenosky 1988)。私の研究でも、シロザとシラカシの最大活性の違いはたかだか10%でした。しかしこれまでのところ数少ない測定のほとんどは最大活性のみしか測っておらず、Kc、Koなどといった測定しにくいものになると、報告は皆無に近いのではないかと思います。
話をMakino et al. (1988) に戻すと、結局のところ同じPNUEだからといって上に上げた4つの候補が同じだとは言えないわけで、PNUEの種間差をもたらす要因は、「どういう種を比較するのか」で変わってくる、と考えたほうがいいのかもしれません。
Warren and Adams (2004) は、常緑植物の葉で光合成能力が低いのはRuBPCaseが無駄に投資されているためだ、と主張するレビューを書きました。彼らが行った実験では、20数種のRuBPCase濃度と葉窒素濃度を比較したところ両者の間に相関がなく、RuBPCaseあたりの光合成速度にも大きなばらつきがあり、ある種ではRuBPCaseを無駄に蓄積していることが示唆されました(Warren et al. 2000)。また、マツのある種では過剰なRuBPCaseの蓄積が見られました(Warren and Adams 2001)。彼らは常緑植物ではRuBPCaseが貯蔵タンパク質として蓄積されているためこのようなことが起こると考察しています。私は彼らの結果には多少疑問を持っているのですが、とりあえずこういう議論もある、ということで。
違いがあるのはわかりましたが、そこにどういう意味があるのでしょうか? 最初の疑問に戻ると、なぜわざわざ低い光合成能力を持っているのでしょうか? また、光合成能力が低いにも関わらず、淘汰されずにいられるのはなぜでしょうか?
今のところ、これらの疑問に答えるための明確な証拠は得られていません。しかし、いろいろ推測することはできます。
おそらく、低い光合成能力を持つ種は、なんらかの性質を有利にするための犠牲として光合成能力が低くなっているのだと思われます。傍証としてよく知られているのが、LMAが高い種では葉の物理的強度が高い、ということです(Reich et al. 1991)。葉の物理的強度を高めるためには葉肉細胞の細胞壁を厚くすることが必要で、その結果葉の中に二酸化炭素が拡散しにくくなっているかもしれません。実際、我々は葉肉細胞の細胞壁の厚さとRuBPCaseあたりの光合成速度の間に負の相関があることを示しています(Terashima et al. 1995)。
多くの研究で共通してみられた「光合成系への葉窒素分配の違い」は、光合成系以外への窒素投資が重要なのではないでしょうか。葉っぱを丈夫にするためには、ある程度窒素も必要だとか、あるいは、葉の寿命を長くするためには、窒素を含んだ二次代謝化合物を多く持つ必要があるのかもしれません。
多くの研究で共通してみられた「光合成系への葉窒素分配の違い」は、光合成系以外への窒素投資が重要なのではないでしょうか。葉っぱを丈夫にするためには、ある程度窒素も必要だとか、あるいは、葉の寿命を長くするためには、窒素を含んだ二次代謝化合物を多く持つ必要があるのかもしれません。我々は細胞壁の窒素に着目し、PNUEが低い葉では細胞壁への窒素投資が多いことを発芽時期が異なるイタドリ(Onoda et al. 2004)とコナラ属の常緑・落葉種間(Takashima et al. 2004)で発見しました。このことから、葉の寿命が長い植物では光合成への窒素投資を犠牲にして細胞壁を増やし、葉を丈夫にしているという、光合成と寿命の間のトレードオフがあることを示唆しました。