光合成の生理生態学講座
目次
はじめに (200512 01)
直観的な説明 (200512 01)
数式を使った説明 (200512 12)
RuBPが飽和しているときの光合成速度 (200512 12)
RuBPが足りないときの光合成速度 (200512 18)
とどのつまり (200512 23)
リン酸律速 (200512 23・200512 28)
光合成速度が違う、ということをこのモデル上で考えると? (200512 28)
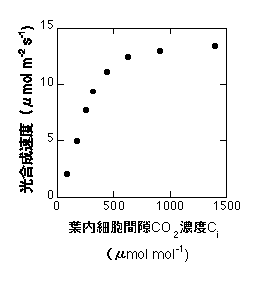
現象としては、二酸化炭素濃度が高いほど光合成速度は高くなります。飽和光下で光合成速度のCO2依存性は以下のようになります。
ここでは、反応速度論に基づいたモデルを紹介することを通して、光合成速度とCO2濃度の関係がどのように決まっているかを理解します。このモデルは光合成の生化学をもとにしていますので、このモデルを理解することがそのまま光合成速度がが決まる生化学的基礎を学ぶことになります。
ここで紹介するモデルはFarquharらによって作られました(Farquhar et al. 1980, Farquhar and von Caemmerer 1982, von Caemmerer 2000)。このモデルにより光合成の環境応答の理解は飛躍的に深まり、以後現在に至るまで、このモデルの存在意義は衰えることを知らず、この分野の「常識」と化しています。これから光合成を環境応答の面から研究しようとしている人は、是非このページだけでもプリントアウトして、理解していただきたいと思います。
Farquhar らのモデルは、残念ながら単純なものではありません。ここでは、このモデルの本質を理解していただくために、まずCO2濃度と光合成速度の関係を直観的に説明し、そのあとで数式を使った説明をすることにします。
光合成の話をする前に、酵素反応における反応速度と基質の関係の基礎から簡単に説明します。一般的に、反応速度は基質の濃度に依存します。基質の濃度が低いときには、基質の濃度が高くなるにつれ、反応速度も直線的に増加なりますが、基質濃度の増加にしたがって反応速度の増加は鈍くなり、ある一定の速度を上回ることはなくなります(飽和)。基質濃度が高いほど反応速度が高くなるわけは、濃度が高いほど基質と酵素が出会う確率が高い(出会わないと反応が起こらない)、と考えるとよいでしょう。
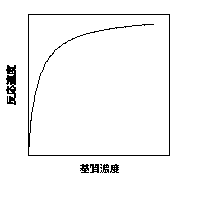
反応速度Vと基質濃度Sの関係の典型。直角双曲線V = Vmax S/(S + Km)で表される。発見者にちなみミカエリス-メンテン式と呼ばれる。
Farquharのモデルは、この反応速度論をもとに考えています。光合成の場合は、CO2が基質でRuBPCaseが触媒する酵素と考えることができます。しかし、CO2と光合成速度の関係は上記のような単純な直角双曲線で表すことはできません。これは、RuBPCaseが二つの反応を触媒するからです。一つは、1分子ずつのRuBPとCO2を結合させ、2分子のPGAを作る反応です(カルボキシル化反応)。もう一つは1分子ずつのRuBPとO2を結合させ、PGAとPGlAを1分子ずつ作る反応(酸素化反応)です。酸素化が一度起こると、光呼吸で0.5個のCO2が放出されます。というわけで、ここではRuBP、CO2、O2という3つの基質を考えることになります。
RuBPCaseはまずRuBPと結合します。RuBPCase-RuBP複合体にCO2が結合すればカルボキシル化が、O2が結合すれば酸素化が起こります。カルボキシル化が起こる部位と酸素化が起こる部位は同じであるため、O2とCO2は競合する関係にあります(競合阻害)。したがって、O2濃度が上がるとCO2を吸収する速度は下がり、逆にCO2濃度が上がるとO2を吸収する速度が下がります。
RuBPCaseが触媒する反応の速度は、まずRuBP濃度の高低の影響を受けます。RuBP濃度が低い場合、RuBPCaseが触媒する反応の速度はRuBP濃度に強く依存します。これは言い換えると光合成の律速段階がRuBPCase触媒反応ではなく、その上流(例えば電子伝達系やカルビンサイクルの別の酵素など)であることを意味します。ここではまとめて「RuBP再生系」と呼ぶことにしましょう。RuBP再生系の能力が高い場合、つまりRuBP濃度が高い場合は、光合成速度の律速段階がRuBPCase触媒反応(カルボキシル化)になります。
RuBPCaseとRuBP再生のどちらが律速段階になるかは、環境条件によって異なります。RuBPCaseに供給されるCO2が少ない場合、つまりCO2濃度が低い場合はRuBPCaseが律速段階になりやすく、CO2濃度が高い場合はRuBP再生が律速段階になりやすくなります。光環境の変化も影響します。弱光環境では、光化学反応速度が遅く、ATP・NADPHの供給が遅いためRuBP再生速度が光合成速度を律速します。
Farquharらのモデルでは、RuBP濃度が飽和しているか否かで別の数式を使っています。それに倣い、以下ではRuBP濃度が飽和しているか否かで、何が起こっているかを別々に説明しましょう。
最初に、RuBP濃度が飽和しているときのことを考えてみましょう。光強度が高く・CO2濃度が低い状況です。まず一番簡単な話として、O2濃度がゼロの条件の話をします。O2濃度がゼロのとき、カルボキシル化の競合阻害は起こりません。したがってCO2濃度とカルボキシル化速度(=CO2吸収速度)の関係は単純なミカエリス-メンテン式にしたがいます。
しかし、自然界、植物が生きているような条件でO2濃度がゼロという条件はあまりありません(冠水状態などの例外がある)。次は通常のO2濃度での話をすることにしましょう。まずO2濃度が一定であると考えましょう(高等植物では、自然界においてO2濃度が変化する、という条件は想定しにくい)。あと、ミトコンドリア呼吸を無視します。
CO2濃度がゼロのときには、CO2がないのでカルボキシル化が起こりません。このためRuBPCaseは酸素化ばかりを触媒します。このときはもっぱら光呼吸によるCO2放出のみが起こります。
CO2濃度が上昇すると、CO2放出速度が下がり、あるCO2濃度でCO2の出入りがなくなります。このCO2濃度がCO2補償点と呼ばれます。ここではカルボキシル化によるCO2吸収と光呼吸(とミトコンドリア呼吸)によるCO2放出がつりあっています。さらにCO2濃度が上がるとCO2吸収速度(光合成速度)が上がります。RuBP濃度が飽和しているときのCO2吸収速度の上昇には、直観的に言うと三つの意味があります。一つは、一般的な基質と反応速度の関係として、基質が増えるほど反応速度が上がることです(イ)。二つめは、CO2濃度がO2濃度に対して相対的に増えることにより、RuBPがカルボキシル化によって消費されるか酸素化によって消費されるかの割合が変わり、カルボキシル化の割合が上がることです(ロ)。三つめは、酸素化の割合が減り、光呼吸によるCO2放出速度が低下することです(ハ)。CO2吸収速度の急激な増加のほとんどはイで説明できます(あくまで直観的な説明。実際にはイとロの二つは区別できない)。
次に、RuBP濃度が飽和していない状況を考えましょう。CO2濃度がかなり高くなります。RuBPが一定の速度で供給されているとすると、RuBPCaseがRuBPに結合する速度はRuBPが供給される速度によって規定され、一定になり、CO2濃度に依存しません。しかしこのときCO2吸収速度はCO2濃度によって変化します。これは、CO2濃度によってカルボキシル化と酸素化の速度比が変化するためです。つまりこの状態ではロとハの影響によって光合成速度が変わっているわけです。光合成速度はCO2濃度上昇とともにゆるやかに増加します。
他に、「リン酸律速」とよばれる別の律速様式もありますが、それは後ほど別に述べます。
では数式を使いましょう。理解しにくくなりますが、正確になります。都合上数式を一行で表しているので、分数がわかりにくくなっていると思いますが、ご了承下さい。
まずは葉のCO2吸収速度です。葉では、カルボキシル化によるCO2吸収、光呼吸によるCO2放出、ミトコンドリア呼吸のCO2放出の和となります。カルボキシル化速度をVc、酸素化速度をVoとすると、CO2吸収速度Pは以下の式になります。
P = Vc - 0.5Vo - Rd (1)
Rdはミトコンドリア呼吸です。光があたっているときのミトコンドリア呼吸速度は暗黒時の呼吸速度(暗呼吸速度)とは違うので、day respirationと呼ばれます。Voに0.5がかかっているのは、酸素化一度あたりCO2が0.5出ていくことを意味します。Vcは RuBPが飽和しているときとそうでないときとで式が異なります。
光合成の前に、ミカエリス-メンテン式から行きましょう。上で述べたように、基質濃度Sと反応速度Vの関係は直角双曲線(ミカエリス-メンテン式)で表すことができます。
V = Vmax S/(S + Km) (2)
ミカエリス-メンテン式は、二つの定数Vmax・Kmで表されます。Vmaxは基質濃度が充分高いときの最大反応速度です。KmはVmaxの半分の速度を実現するような基質濃度で、ミカエリス-メンテン定数と呼ばれます。 おおざっぱに言うと、Vmaxは基質濃度が低いときの速度、Kmは基質濃度が高いときの速度に影響します。
上述したように、RuBPCaseが触媒する反応にはRuBP、CO2、O2の三つの基質が存在します。このため上のような単純な式ではなく、もっと複雑な式になります。ただし、RuBPが飽和しているときには、RuBPの濃度の影響を無視することができます(飽和=濃度が変化しても速度が変わらない)。強光・低CO2濃度ではRuBP濃度は飽和しやすくなります。
RuBP濃度が飽和しているとき、Vcは以下の式で記述されます。
Vc = Vcmax C/{C+Kc(1+O/Ko)} (3)
ここで、CとOはそれぞれCO2とO2の濃度です。KcとKoはそれぞれCO2、O2にたいするミカエリス定数で、Vcmaxは最大カルボキシル化速度です。この式は、競合阻害時の一般的な反応速度式です。よく見るとミカエリス-メンテン式によく似ています。違うのはKmの部分です。ミカエリス-メンテン式でのKmは、式3ではKc(1+O/Ko)になっています。このことは、式3は「酸素濃度(O)が変化するにしたがってKmが変わるミカエリス-メンテン式」であるということを意味します。Oがゼロならば式3は式2と全く同じです。Cが一定の場合、Oが増大するとKmが増加し、Vcが低下します。一方、Oが一定ならば、VcはCに依存して増加します。競合阻害式について詳しく知りたい方は、お手元の生化学の教科書を見て下さい。反応速度論を説明しているところに必ず書いてあります。
同様に、Voも表すことができます。
Vo = Vomax O/{O+Ko(1+C/Kc)} (4)
Vomaxは最大酸素化速度です。
式4を式3で割るとVcとVoの比Zを示すことができます。
Z = Vo/Vc = (Vomax O Kc) / (Vcmax C Ko) (5)
また、2Vc = Vo となるCO2濃度、つまり、光呼吸のCO2放出とCO2吸収速度が釣り合うことにより、呼吸速度を無視したときにCO2吸収速度が0になるCO2濃度(G*)が以下の式で求められます。
G* = ZC/2 = (Kc O Vomax) / (2Ko Vcmax) (6)
このG*は実測が可能なため、Zよりも便利なパラメータとしてよく利用されます(こちら)。また、RuBPCaseのキネティクスの性質を一つの数値で表現するためにspecificity factor(t)というパラメータが利用されます。
t = (Ko Vc)/(Kc Vo) = (Ko Vcmax)/(Kc Vomax) = 0.5 O/G* (7)
式3-6を使うと、RuBPが飽和しているときの光合成速度Pcを以下のように表すことができます。
Pc = Vc (1 - 0.5Z) - Rd
= Vc (1 - G*/C) - Rd
= Vcmax (C - G*) / {C+Kc(1+O/Ko)} - Rd (8)
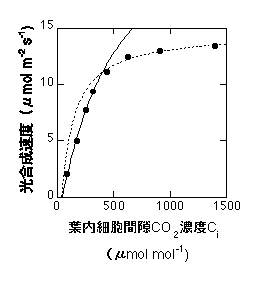
この式の(C - G*)のうち、Cの項(正確にはVcmax C / {C+Kc(1+O/Ko)})がcarboxylation速度を、G*の項が光呼吸によるCO2放出を表すことになります。G*は普通のO2濃度(21%)のときに約40 ppm(0.004%)です。C3植物間ではそれほど大きな種間差はありません(Brooks & Farquhar 1985など。ただし、全く同じというわけではありません)。以下は式8の適用例です。低CO2濃度でのみ合います。Vcmaxは高いので飽和点は見えません。
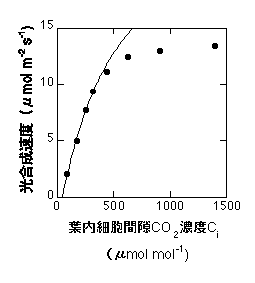
式8の適用例。
一定の温度ではKc、Ko、 G*は基本的に定数で、種間差も大きくありません。しかし、葉に含まれるRuBPCaseの含量が葉によって大きく異なるため、Vcmaxは葉によって大きく異なります。逆に言うと、RuBPCaseを多く持った葉は高いVcmaxを持ち、高い光合成速度を持つことになります。同一種内では多くの場合VcmaxとRuBPCaseの間にきれいな相関が見られます(例えばvon Caemmerer and Farquhar 1981)。
RuBPが足りないときは、RuBPの供給速度が光合成速度に最も大きな影響を与える要因となります。Farquharらのモデルでは、RuBPの再生速度がチラコイド膜の電子伝達によって律速されていると考え、電子伝達速度を重要なパラメータと位置づけています。まずは、電子伝達速度とカルビンサイクル・光呼吸の関係を見ましょう。
カルビンサイクルでは、1分子のPGAを還元するのに1モルのNADPHを必要とします。1モルのNADPHを電子伝達において還元するためには2モルの電子を伝達する必要があります。1モルのRuBPのカルボキシル化では2モルのPGAが合成されますから、カルボキシル化あたり4電子が必要です。
光呼吸では、1モルのRuBPを酸素化すると1モルずつのPGAとPGIAが合成されます。1モルのPGIAからは0.5モルのPGAが合成されます。光呼吸の過程では、1モルのPGlAから0.5モルのアンモニアが放出されますが、このアンモニアは1モルの電子を使って最固定されます。したがって、1モルのRuBPを酸素化するごとにPGlAに1モル、PGAに3モル、合計4モルの電子伝達を必要とします。
カルビンサイクルと光呼吸を合わせると、電子伝達速度J=4Vc+4Voということになります。ここでZ(=Vo/Vc)を導入すると、カルボキシル化を一度行うためには、4+4Z個の電子が伝達されなければいけない、ということになります。
J = Vc (4+4Z) (9)
したがって、RuBP量が律速しているときの光合成速度Prは以下のように表されます。 式1に式9を代入します。
Pr = Vc (1 - 0.5Z) - Rd
= (1 - G*/C) Vc - Rd
= (1 - G*/C) J /(4+4Z) - Rd
= J (C - G*) /(4C+8G*) - Rd (10)
式10は以下のようになります。高CO2領域のみで合います。
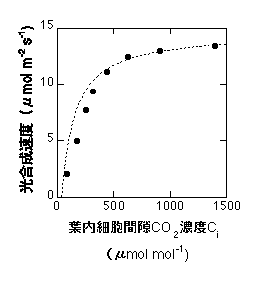
10式の適用例。
式9、10では、RuBP生産速度がNADPH生産に律速されていると考えていますが、Farquhar & von Caemmerer (1982)ではATP生産に律速される可能性も考えています。詳細は省きますが、このときは以下の式で表されます。
Pr= J (C - G*) /(4.5C+10.5G*) - Rd (11)
考慮に入れるのがATPとNADPHで式が違ってしまうのは、電子伝達あたりで生産されるNADPHとATPの比と、光合成において消費されるNADPHとATPの比が異なることに由来します。ただ、こちらでも書きましたが、一度の電子伝達でいくつATPが生産されるのかはまだ確定していません。80年代は一度のATP生産のために3プロトンの輸送が必要である、という説が広く信じられていたように思いますが、私が知る最新の研究では4プロトンになっています。また、電子伝達速度とATPの生産速度は一定ではないことがわかってきています(詳しくは「光阻害」で書きます)。式11は80年代はよく使われましたが、現在の知見からするともう正しいものではないと思われますし、実際式11が使用されている論文を見ることも少なくなりました。ただし、電子伝達・ATP生産・RuBP生産の関係は確定しているわけではないので、将来どうなるかはわかりません。
さて、電子伝達速度Jがどのように決まるかです。Jは、RuBP生産速度が光合成速度を律速する条件では、光強度と温度によって変化します。CO2濃度には依存しないと考えられています(RuBP生産以外が光合成を律速する場合は、他の反応段階の速度変化に合わせてJも変化する。その変化は式10にしたがう)。温度については「温度と光合成」で書きます。
光強度が低くなれば、光化学反応が電子伝達速度を律速します。このため電子伝達速度は光強度に対して飽和型の曲線を示します。詳しくは「葉光合成のモデリング」で説明する予定ですが、様々な研究者が様々な式を使ってモデル化しています。ここでは、非直角双曲線を使っておきます。
J =(fr I + Jmax - {(fr I + Jmax)2 - 4fr I qr Jmax}0.5)/(2qr) (12)
ここで、frは初期勾配、Iは光強度、Jmaxは光飽和時の最大電子伝達速度、qrはカーブの凸度です。詳しい説明は「葉光合成のモデリング」で行います。
JmaxはRuBP再生にかかわる多くのタンパク質量・活性と相関を示します(例えば von Caemmerer and Farquhar 1981)。Jmaxを律速している反応段階が何かはわかっていません。電子伝達活性という説が有力ですが、FBPaseという説も過去にはありました。光合成系にかかわる多くのタンパク質の量がJmaxと比例しているためどこが律速段階かは相関関係からはわかりません。また、電子伝達活性がJmaxを決めていたとして、電子伝達系のどこが律速段階かも、明らかではありません。プラストキノンの電子運搬が有力視されています。
葉が実現する光合成速度は、RuBPが律速しているときの光合成速度PrとRuBPが飽和しているときの光合成速度Pcの「低いほう」として表されます。
P = min{Pr, Pc} (13)
式をあてはめた例。
ただし式13はあくまで単純化したもので、PrとPcの値が近い場合は、理論上複雑なものになります。例えば、CO2を徐々に上げていくと、律速段階がPcからPrにシフトするのですが、実際には式13や上図で表されるようにある点できれいにシフトするわけではなく、もう少しゆるやか、つまりどこが境目かがはっきりしないような変化をします。これにはいくつか理由があります。一つは葉緑体中のRuBP濃度です。式8では何度か説明したようにRuBP濃度は飽和していることが仮定されています。一方式11では、Jが一定ならばRuBP濃度は一定になると考えられます(説明はしていませんが)。つまり式13では、あるCO2濃度でRuBP濃度が飽和から突然ある濃度(Xとしましょう)まで変化することを仮定しているわけです。実際にはRuBP濃度は飽和レベルからXまで徐々に変化しているわけで、そのときの速度を記述するには式8や式11より複雑な式が必要です。ただ、RuBP濃度が飽和レベルからXまで変化する範囲はかなりせまいことが、理論的には期待されています(RuBPに対するRuBPCaseのKm値が低いからなのですが、詳しい説明は省きます)。もう一つの理由は、葉緑体によって性質が若干異なるため、全ての葉緑体が一斉にPcからPrにシフトすることはなく、葉全体ではゆるやかにシフトしているように見えるということです。しかしこれらの影響は小さく、上図で見られるように、実測データを説明するには式13で充分です。
Farquharのモデル(Farquhar et al. 1980)から5年後、第3の律速段階、リン酸律速を取り入れた改良モデルが提案されます。Sharkey (1985) です。
リン酸律速とは、葉緑体内の無機リン酸の濃度が光合成速度を律速する、というものです。トリーオスリン酸律速とも呼ばれます。光合成産物は葉緑体から細胞質へ輸送されるわけですが、このときの糖は3つのCと1つのPを持ち、トリオースリン酸(triose-phosphate)と呼ばれます。このトリオースリン酸からショ糖が細胞質内で合成されるわけですが、その過程でリン酸が放出され葉緑体へ戻ります。細胞質から葉緑体内へ入るとき、リン酸は包膜の輸送タンパク質(リン酸トランスロケーター)を通過するのですが、リン酸はトリオースリン酸との1対1の交換で葉緑体に戻ります。つまり、トリオースリン酸が出ていくときにはリン酸が戻ることが不可欠で、逆もまたしかりです。戻ったリン酸はチラコイド膜でADPと結合してATPとなり、再びカルビンサイクルに入ります。光合成速度が増加し、細胞外ショ糖の処理能力を上回る(他の器官でショ糖を利用しきれなくなる、あるいは師管輸送能力を越える)ときにこのリン酸律速が起こります。ショ糖を処理しきれないため、細胞質内にショ糖が蓄積していきます。このとき、細胞質内に糖リン酸も蓄積していきます。すると、無機リン酸が放出されないので細胞質内の無機リン酸濃度が下がります。結果的に、リン酸が不足するとトリオースリン酸が出て行かなくなるし、ATPの合成もできなくなります。この結果、光合成速度はトリオースリン酸とリン酸の代謝回転速度に依存することになります。このときの光合成速度Ppは以下の式で表されます。
Pp = 3T - Rd (14)
ここで、Tはトリオースリン酸が葉緑体から出ていく速度です (Sharkey 1985)。
とっても単純な式です。Tは温度が一定ならば変化することはなく、リン酸律速が起こっているときには、光合成速度はCO2濃度、O2濃度に依存しません。リン酸律速が起こっていることは、普通は光合成速度にCO2濃度依存性がなくなることで知ることができます。ただし、RuBP律速のときもCO2濃度依存性は小さいので、厳密にはO2濃度を変えても光合成速度が変わらないことを確認する必要があります。
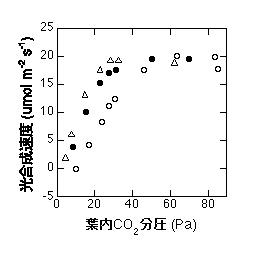
セイヨウキョウチクトウでO2・CO2濃度を変えて測定した光合成速度。O2濃度は三角で10%、黒丸で20%、白丸で50%。CO2分圧50Pa(500ppm)以上で光合成速度に違いがないので、リン酸律速が起こっていると考えられる。光合成速度は20が上限のようですから、呼吸速度を1とすると、T=21/3=7と考えられる。
かつてO2濃度依存性は光呼吸の影響を知るために調べられていました。光呼吸はO2濃度が高ければ起き、低ければ起きないと考えられていたためです。上の図のように、リン酸律速が起こっているときにはO2濃度依存性が見られなくなります。しかしここから「光呼吸が起こっていない」と結論するのは間違いです。実のところ、光呼吸はO2濃度が下がると起こりにくくなるのですが、カルボキシル化も起こりにくくなっているために、見かけ上CO2吸収速度が変わらないのです。これは光合成が下流(糖輸送)によって律速されているためです。条件がどうあれ、トリオースリン酸/リン酸の輸送速度に限度がある場合は、その限度を超えてCO2が葉に入ってくることができないのです。このあたりはいくら言葉を使って説明しても理解するのは難しいかもしれません。詳しく勉強したい方はSage (1990) をどうぞ。
私の経験では、普通に植物を育てているときには、大気CO2濃度でリン酸律速が起こったことはありません。かなり高CO2濃度で光合成速度を測定すると見られることがあります。また、トリオースリン酸の利用速度は温度依存性が強い(低温と高温での速度の違いが大きい)ことが知られています。そのため、普通の温度で育てた植物の光合成速度を低温で測定すると大気CO2濃度でも光合成速度がリン酸律速になることがしばしばあります(Labate & Leegood 1988, Sage 1990, Cen and Sage 2005)。
同じ条件で測定しても、光合成速度は葉っぱによって違います。このことはモデルに則って考えると、どう説明されるでしょうか? まず、式8と10で、変化しうるもの(変数)と変わらないもの(定数)が何かを考えてみましょう。
式8では多くのパラメータが出ていますが、環境因子としてはOとCがあります。これは同じ条件にした、と考えましょう。あと、KoとKcがあります。これはミカエリス定数と呼ばれているように、定数です。種による違いも小さいと考えていいでしょう。G*も種によってあまり変わりませんので、定数と考えていいでしょう(これらのパラメータは温度によって変化しますが、ここでは無視します)。そうすると、残りはVcmaxになります。ここまで、Vcmaxの単位を説明してきませんでした。Farquharのモデルは酵素あたりでも葉緑体あたりでも適用できますが、ここでは葉レベルで考えてることにしましょう。つまり、葉面積あたりの最大酵素活性がVcmaxである、ということになります。Vcmaxが葉っぱごとに違う場合、考えられるのは、葉っぱにどれだけ酵素が存在しているかが異なることです(他にも、酵素の性質が違う、活性化レベルが違うなどの可能性があります)。したがって、RuBPが飽和しているような条件で光合成速度が違った場合、酵素RuBPCaseの量が違うことが原因と考えられます。
式10では、変数はJとCだけです。Jは式8で与えられます。まず、光が飽和しているとしましょう(I →∞)。そうすると、J≒Jmaxになります(数式については「光合成のモデリング」で説明しますので、わからない方は、「そういうものなのだ」と考えて下さい)。Vcmaxの議論同様、Jmaxの違いが光合成速度の違いになるわけです。Jmaxは最大電子伝達速度ですので、電子伝達能力が高い葉ほど強光・高CO2濃度での光合成速度が高くなります。
VcmaxとJmaxの値は葉によって大きく違います。しかし、両者の比(Jmax/Vcmax比)は種・葉の間ではそれほど大きくは変化しません。だいたい、通常CO2濃度前後(300~500ppm)でPrとPcの値が同じ程度になるようなバランスを保っています。
次に光が弱い場合についてですが、光強度が非常に弱い場合、Jはfrで決まります(と考えて下さい)。「光と光合成」で光-光合成曲線の初期勾配fについてお話ししましたが、frは光-電子伝達速度の初期勾配です。光-光合成曲線について説明した理屈とほとんど同じ理屈で、クロロフィル含量が高いほどfrが高くなります。
Cを一定にする、という仮定で話をしてきましたが、葉っぱの周りの空気のCO2濃度が同じだからといって、葉の中の細胞間隙のCO2濃度や、葉緑体の中のCO2濃度が同じだとは実は限りません。特に、前者は気孔の開き方によって、様々に変化し得ます。Cがどのように決まるかは「二酸化炭素の拡散」で説明します。