光合成の生理生態学講座
目次
はじめに (200511 09)
暗呼吸速度(光強度が0のときの光合成速度) (200511 09)
初期勾配(弱光下の光合成速度) (200511 09)
光−光合成曲線の凸度(中くらいの光での光合成速度) (200511 12)
最大光合成速度(強光下の光合成速度) (200511 12)
光照射下の暗呼吸速度 (200511 12)
光は光合成におけるエネルギー源です。当然、光がなければ光合成は起こりません。弱光下では光合成速度は低く、強光下では光合成速度は高くなります。しかし、光強度が高くなると光合成速度はある一定以上にはならなくなります。ここでは、光−光合成曲線の形を通して光強度がどのように光合成速度を調節しているかを考えます。
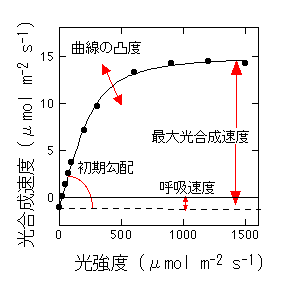
下に典型的な光−光合成曲線を示します。
シロザの光−光合成曲線を非直角双曲線(P =(fI+ Pmax - {(fI+ Pmax)2 - 4fIqPmax}0.5)/2q - R)で回帰したもの。式については下参照。
条件によって、あるいは葉によって光強度と光合成速度の関係は異なります。これを定量的に比較するためには、曲線の特徴づけが必要です。最も合理的なのは数式で回帰し、その係数の違いを比較することです。光−光合成曲線を回帰する数式について詳しくは「葉光合成速度のモデリング」で紹介します。ここでは最もよく使われる非直角双曲線で近似したものを示しています。非直角双曲線は4つの係数で構成されています。呼吸速度、初期勾配、曲線の凸度、最大光合成速度です。この項ではこれら4つの係数がどのように決まっているかを説明します。
光がない状態では光合成は行われず、暗呼吸による二酸化炭素を放出のみが起こっています。
暗呼吸速度がどのように決まっているかは複雑で、不明な点も数多くあります。いずれ機会があれば詳しく書きますが、ここでは簡単な紹介に留めます。暗呼吸速度は環境要因だけではなく、細胞内の生化学的な状況にも依存します。多くの植物では呼吸の基質である糖の濃度が高いときに呼吸速度が高くなります。一般に、陽が沈み夜が始まった頃には、昼間の光合成によって生産された糖がまだ葉に蓄積しており、呼吸速度が高いです。時間が経つと、糖の含量は転流と呼吸消費によって徐々に低下するため、呼吸速度も低くなります(Azcon-Bieto et al. 1983, Azcon-Bieto and Osmond 1983, Noguchi et al. 1996)。近年、陰生植物クワズイモでは呼吸速度が糖濃度に依存せず、呼吸速度の日変化も示さないことが示されました(Noguchi et al. 1996)。このような植物では、呼吸速度は細胞内のエネルギーレベル(ADP/ATP比)に依存します。
暗呼吸速度は葉によって異なり、最大光合成速度と相関があることが知られています。気温25度前後で、だいたい最大光合成速度の1/20-1/10といった値です。この違いは暗呼吸にかかわるタンパク質の量に依存するのではないかと私は思っていますが、実際のところどうだかは知りません。
光照射下では呼吸速度は低くなります。詳しくは下をご覧下さい。
光強度が100 μmol quanta m-2 s-1以下では、光強度と光合成速度の関係はほぼ直線です(ただし、後述するようにKok効果というのがあり、必ず直線になるわけではないことに注意が必要です)。下は私がセイヨウキョウチクトウで測定した例です。
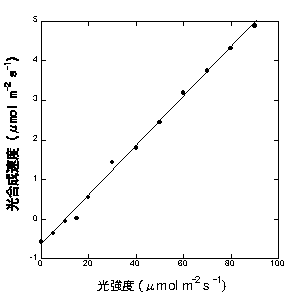
測定条件は通常CO2濃度・葉温25度(詳細忘却)
光合成速度が0になるような光強度を光補償点とよびます。教科書ではよく出てきますが、きちんと測定するのはめんどうなこともあり、論文ではあまり使わない指標です。そこで直線部分について光強度と光合成速度の直線回帰をします。そうすると、回帰式(Y = a + bX)のY切片(a)が呼吸速度で、傾き(b)が、あたった光あたりの光合成量ということになります。後者を、「光−光合成曲線の初期勾配」と呼びます(みかけの量子収率といったりもします)。この例ではY=0.0624X-0.66ということで、呼吸速度は0.66 μmol CO2 m-2 s-1、初期勾配が0.0624 mol CO2 mol-1 quantaということになります。ここでは、この初期勾配がどう決まるのかについて書くことにします。
初期勾配の数値(ここでは0.0624 mol mol-1)には様々な意味があります。この数値は弱光下での光合成速度の指標となるだけでなく、光合成の光利用効率も表します。この図では横軸の光強度がモル光量子で、縦軸の光合成速度がモルCO2で表されていますから、傾きは光合成における「葉にあたった光量子数あたりの吸収CO2」を示すことになります。この場合は、1光量子あたり0.0624CO2を吸収できる、ということになります。
初期勾配は「あたった光あたりの光合成量」です。初期勾配を高くするには、あたった光をたくさん吸収するか、吸収した光を効率よく光合成に利用するかのどちらかの方法があります。
まず光吸収について考えましょう。葉は受けた光を全て吸収できるわけではありません。多くの場合、受けた光のうち5-10%は反射され、5-10%は透過してしまいます。したがって残りの80-90%が吸収された光ということになります(光吸収率)。光吸収率の大小に最も重要なのは葉のクロロフィル含量です。クロロフィルが多ければ多いほど葉が光を吸収しきるチャンスが多く、吸収率が高くなります。
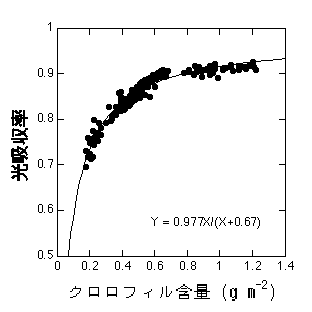
シロザにおけるクロロフィル含量と光吸収率の関係。加藤真晴氏測定。
このデータは一年草のシロザのものですが、多くの草本植物は同様の曲線にのると考えていいと思います(Evans 1989)。しかし、葉の表面にワックスが多い常緑木本植物などはかならずしもこの線にのらないようです(Syversten et al. 1995)。つまり、同じクロロフィル量を持つからといって同じ吸収率を持つとは限らないようです。
次に、吸収した光の利用について考えましょう。では、吸収した光でどれだけ光合成ができるでしょうか? 吸収光量あたりのCO2吸収量(もしくはO2発生量)を量子収率(quantum yield)と言います。量子収率が生化学的にどのように決まっているかはこちらに書きました。実現可能な最大量子収率は0.125 mol CO2 mol-1 quantaということになります。しかし、上の図もそうですが、実際の葉でこのような値になることはありません。現実の量子収率が理想よりも低くなる理由はいくつかあるのですが、最も大きな理由は光呼吸の影響です。こちらに書きましたように、光呼吸では酸素化が起こるたびにエネルギーが消費され、CO2が失われます。このエネルギー消費・CO2損失の両方が見かけ上量子収率を低下させます。通常条件での低下は30-50%程度です(光呼吸の影響について詳しくは「二酸化炭素と光合成」で書きます)。また、光呼吸の影響は温度によって変化します。光呼吸は相対的に高温で起こりやすいため、初期勾配は高温で低下します(詳しくは「温度と光合成」で書きます)。光呼吸はCO2濃度を上げる、あるいはO2濃度を下げることによって抑制することができます。このような条件では、多くのC3植物で量子収率は0.10から0.11という理想値にかなり近い値になります(Bjorkman and Demmig 1987)。
他にも量子収率を低下させる要因はあります。一つは光合成系がもつ本質的なムダです(こちら)。あとは、クロロフィル以外の物質が光を吸収した場合、それは光合成に利用されませんので量子収率が低下します。また、量子収率は光化学系IIに傷害が起こると低下します。これは、傷害を受けた光化学系IIに結合したクロロフィルが光を吸収しても、そのエネルギーが光合成に利用されないためです。量子収率と損傷を受けていない光化学系IIの割合は比例します(例えばBjorkman and Demmig 1987)。
以上の要因から、初期勾配の値は、健康な葉を使って通常CO2濃度、通常温度で測定した場合、0.04から0.06 mol CO2 mol-1 quantaという値になります。私の経験では、初期勾配の誤差の原因となりやすいのは、光合成速度の測定よりも光強度の測定誤差です。光合成速度に使うCO2濃度を測定する機械(赤外線ガス分析機)は、標準ガスを購入すれば比較的楽に校正ができますが、光センサの校正は素人にはできません。また、光合成測定時に葉に均一な光強度を当てるのもけっこう難しい技術です。あとは光の質の問題があります。上の例はライカ社のLi-6400という光合成測定装置で測定したものですが、この装置の光源はLEDで、赤い光と青い光が使われています。クロロフィルは青と赤を吸収しやすいので、太陽光で測定するよりも吸収率が高く、初期勾配も高くなるはずです。上の図の初期勾配が高めなのもそのせいだろうと思います。
光−光合成曲線の形は、初期勾配(弱光域の直線部分の傾き)と最大光合成速度で多くの場合代表できます。しかしその間の形はどの葉でも同じ、というわけではありません。中間の部分をどのように数値化するかは難しいのですが、曲線の「凸度」として評価されることが多いです。一般に、光−光合成曲線は以下のような非直角双曲線で近似されます。
P =(fI+ Pmax - {(fI+ Pmax)2 - 4fIqPmax}0.5)/2q - R
ここで、fが初期勾配、Iが光強度、Pmaxが最大光合成速度、qが曲線の凸度(0<q<1)、Rが呼吸速度です。このqが小さいほど曲線は直角双曲線に近づき、大きいほど二直線に近づきます(下図・「光合成のモデリング」参照)。
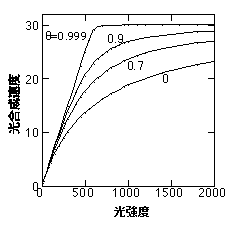
非直角双曲線。Pmax=30、f=0.045、R=0を仮定している。
ここでは、このqがどのように決まるかについて書きます。
この凸度がどのように決まっているかについては、いくつか説が提唱されています。古くは葉の中のCO2の拡散と固定のバランスでqが決まると考えた人もいましたが(Rabinowitch 1951)、たぶん正しくないでしょう。現在、二つの重要な説があります。順番に紹介しましょう。
まず憶えていただきたいのは、葉の光−光合成曲線は、葉の表から光を当てたときと、裏から光を当てたときで異なる、ということです(Terashima & Saeki 1985, Terashima 1986)。生データは持っていないので、イメージ図をどうぞ。
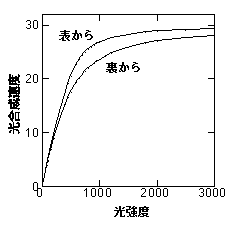
言ってしまうとこの図はqを変えただけです。どうして表と裏で曲線の形が違うかを説明すると、qの「成り立ち」が理解してもらえるはずです。ややこしい話ですが、おつきあい下さい。
qの話をする前に、確認として表からでも裏からでも、弱光及び強光での光合成速度は両者でそれほど変わらない、ということを理解してもらいましょう。とても弱い光強度では光合成速度は当たる光の量で決まります。表だろうが裏だろうが吸収された光は光合成に利用されますので、同じ光強度なら同じ量の光合成が行われるわけです(ただし、葉の裏に毛が生えている場合など、表と裏で光の吸収率が変わってしまう場合はこの限りではありません)。一方、とっても強い光が当たると、全ての葉緑体の光合成速度が飽和してしまいます。そのため、表から当てようが、裏から当てようがやっぱり光合成速度は似たような値になります。
では、中くらいの光強度での光合成速度がなぜ違うかを説明しましょう。まず理解してもらわなくてはいけないのは、1)葉の光合成とは、葉緑体の光合成の総和である、ということです。そして、2)各葉緑体の光環境は均一ではありません。表側から光を当てたときには、表側にある葉緑体が強い光を受けます。表側の葉緑体がかなりの光を吸収してしまう結果、裏側にある葉緑体が吸収できる光強度は弱くなります(Terashima & Saeki 1983)。最後に、3)各葉緑体の性質は同じではありません。クロロフィルあたりの光合成速度を比較すると、表側にある葉緑体のほうが、強光下での光合成速度が高いです(Terashima & Inoue 1984)。以上3つのことが重要です(Terashima & Saeki 1985)。
これらのことが、葉の光合成にとってどのような意味を持つでしょうか? 模式的に示してみましょう。ここでは、各葉緑体の光−光合成曲線が二直線から成ると仮定します。そして、表側と裏側では葉緑体の光合成能力が異なります(下図)。
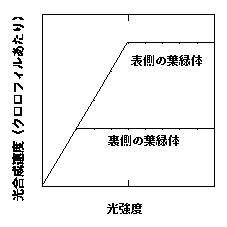
個々の葉緑体の光−光合成曲線の模式図。ここでの横軸光強度は葉緑体が受けた光強度。
葉の表から光を当てて、徐々に光強度を強くしてみましょう。表側の葉緑体は相対的に強い光を受けますが、最初は光が弱いので吸収した光はほとんど光合成に回されます。ある程度光が強くなると、葉緑体の光合成速度は光飽和し、それ以上光合成速度が上がりません。表側の葉緑体は強い光で飽和し、裏側の葉緑体は弱い光で飽和するので、両葉緑体はほぼ同時に飽和するわけです。葉の光合成は、葉緑体の光合成の和です。結果として葉の光−光合成曲線もqが高い二直線になります(下図)。
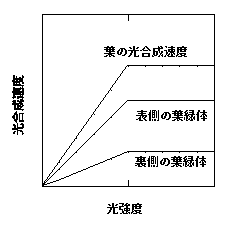
葉の表から光を当てたときの光合成速度。葉の中が二層(表側と裏側)の葉緑体層があることを仮定。葉の光合成速度は二層の葉緑体光合成速度の和となる。横軸は、葉緑体の光強度ではなく、葉の光強度を示している。ある光強度が当たっても、裏側にはあまり光は届かないので、裏側の葉緑体の光合成速度の立ち上がりが鈍い。
今度は、葉の裏から光を当ててみましょう。裏側の葉緑体はすぐに光飽和してしまいます。一方、表側の葉緑体はなかなか光飽和しません。結果として、光−光合成曲線は三直線になります(下図)。qが低いわけです。
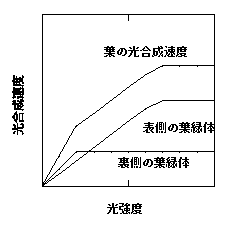
葉の裏から光を当てたときの光合成速度。今度は、表側の光環境が悪いため、表側の葉緑体の光合成速度の立ち上がりが鈍い。
というわけです。重要な点は、「qは、全ての葉緑体が同時に光飽和するような条件で高い」ということです。それぞれの葉緑体が、それぞれの光環境に合わせた光−光合成曲線を持てば良いのです。実は、このような論理展開は、群落光合成における葉の光合成の調節にも当てはまります(「群落光合成」参照)。
ここまでの説明では、各葉緑体の光−光合成曲線は二直線に近い、という仮定を前提にしています。Terashima & Saeki (1985) ではいろいろ文献などで得られた単離葉緑体の光−光合成曲線から計算した値を出して、qが高い、ということを述べています。しかし、Ogren(1993)は異なる光条件で生育した緑藻のデータから、強光で育てた藻類の葉緑体ではqが低いということを報告しています。そこで彼らが提唱したのが次の仮説です。
上記のようにOgren(1993)は強光で育てた藻類の葉緑体ではqが低いことを見出しました。Ogren & Evans (1993) は、葉において、光合成を測定するときのCO2濃度によってqが変わることを報告しています。これらの知見から、彼らはqが低いのは1で紹介したような話とは別で、個々の葉緑体の性質を反映しているのだと考えています。彼らの解釈は以下の通りです。光合成速度は、弱光では電子伝達系の活性によって、強光ではRuBPCaseの活性によって律速されます。電子伝達活性は光が弱いと落ちますが、RuBPCase活性は光強度に依存しません(とします)。ところで、光合成速度は、電子伝達活性に律速されているときも、RuBPCase活性に律速されているときもどちらでもCO2濃度依存性を示します。しかし、その依存性は異なり、CO2濃度が変わったときの光合成速度の変化はRuBPCase活性に律速されているときのほうが大きくなります(詳しくは、「二酸化炭素と光合成」で)。電子伝達活性の光依存性(光−電子伝達速度曲線)はあまり角張っていない(qが低い)と仮定すると、以下のように、光−光合成曲線は低CO2濃度ではqが高く、高CO2濃度では低くなる、ということになります。これは実際の変化をよく説明します(Ogren & Evans 1993, Ogren 1993)。
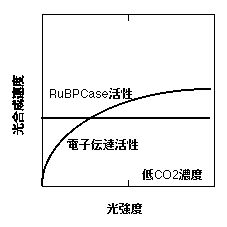
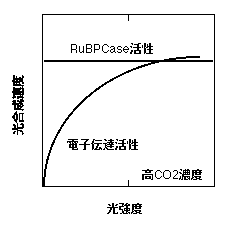
このことから言えるのは、ある葉のqは電子伝達の最大活性(光飽和下の活性)とRuBPCase活性のバランスで決まるということです。RuBPCase活性が相対的に低ければ、qは低くなるわけです。
上記二つの仮説は、どっちが正しくてどっちかが間違っている、という話ではありません。葉の表から光を当てるか裏から光を当てるかの違いは仮説1で説明され、CO2濃度が違う場合は仮説2で説明される、くらいに考えて下さい。ただ、他のケースでどちらがあてはまるかは、ケースによりけり、ということです。では、次に現実のケースを見てみましょう。
qがどういった葉でどういった値を持つのか、また、葉がおかれた環境とどういった関係があるのか、最もよく調べているのは、Leverenzという人です。あいにく私は彼の論文を真面目に読んだことがないので、彼の仕事についてちゃんとしたレビューをすることができません。ただ、たぶん彼は「クロロフィル含量の高い葉ほどqが低い(Leverenz 1987)」と言っているのではないかと思います(すみません。いいかげんで)。
他の文献では、私の知っている範囲では、「窒素含量の高い葉ほどqが低い」です(ただし、相関は低い。Hirose & Werger 1987a, Hikosaka et al. 1999a-生データはこちらを参照)。窒素含量とは相関がない、というデータも出されています(Pons et al. 1989, Schieving et al. 1992)。
これらの結果はどう解釈できるでしょうか? まず、上で述べたような仮説2の葉緑体レベルの違いが考えられますが、どういう葉緑体でqが高いか低いかはまだよくわかっていないので、ここでは「わからない」としかいいようがありませn。
では仮説1はどうでしょうか。よく言われているのが、「葉緑体が弱光に順化しきれない」ということです。葉のクロロフィル含量が多いほどたくさんの光が吸収されます(上参照)。この結果、葉の裏側の葉緑体が受ける光強度はクロロフィル濃度が高いほど低くなります。しかし、どうやら葉緑体は暗すぎる環境には順化できないようで(「光順化」参照)、上で説明したような、「全ての葉緑体が同時に光飽和する」ということができないようです。したがって、クロロフィル含量が高い葉ほどqは低くなってしまうということです。ちなみに窒素含量が高い葉のほうがクロロフィル含量が高い傾向にあり、この考え方と矛盾しません。
裏側の葉緑体が表から光をあてただけでは飽和しない、というのは実測例があります。Green and Kruger (2001) は上から測定チャンバーを工夫することにより、光を上からも下からも当てられるようにし、光合成速度を測定しました。1800μmol m-2 s-1の光をあてたときに比べ、上下両方から1800μmol m-2 s-1の光(合わせて3600)をあてたときにどれだけ光合成速度が促進されたかを比較すると、落葉広葉樹では0-20%程度しか光合成速度が促進されなかったのですが、常緑針葉樹では、多い葉では50%もの促進が見られました。常緑針葉樹では表からかなり強い光を当てても裏側の葉緑体は光飽和しないことが示唆されます。
注意しなければならないのが、qが低くなるのは技術的な問題が原因であるかもしれないということです。育っているときには、水平な葉でも裏から多少は光が当たります。しかし、光合成を測定するときは普通裏から光を当てません(Green and Kruger 2001は例外)。生育時と測定時で葉の中の光環境が違う可能性は高く、この違いがqを低くする可能性はあります。実際、葉の中の葉緑体を調べてみると、最下層の葉緑体は、最下層より一層上の葉緑体よりは明るい環境に順化しているように見えます(たとえばTerashima & Hikosaka 1995)。また、イネ科など、立っている葉では普段から裏にもかなりの光が当たっているはずです。測定された光合成速度と生育条件での光合成速度は、あてた光強度が同じだったとしても、等しいとは限らないと考えたほうがいいでしょう。
強光下では、光合成速度は光強度に依存しなくなります。このことはつまり光以外の要因が光合成速度を律速していることになります。通常大気CO2濃度では、最大光合成速度の律速要因はCO2濃度です。CO2濃度と光合成速度の関係は「二酸化炭素と光合成」で書くことにします。
我々が見ている光合成速度は、厳密には光合成速度から呼吸速度を指し引いたものです。したがって、呼吸速度についても注意を払わないわけにはいきません。
上に、弱光下の光合成速度は光強度に対して直線的に増加する、と書きましたが、厳密には、完全に直線ではなく、光合成速度が0になる光強度(光補償点)を境に傾きが若干変化することが知られています。これをKok効果といいます。
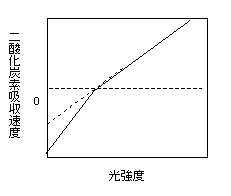
Kok効果のイメージ。ただ、私が測るとあまりKok効果ってはっきり見えないんですよねえ。初期勾配で紹介した図にも全然見られないし。どんだけの頻度で見られるのかとかはよく知りません。
そもそもはクロレラ(緑藻)で発見されたらしいのですが(Kok 1948)、高等植物にも見られます。Kok効果の原因については、光強度が増すとともに、1)暗呼吸速度が低下する、という説と、2)光呼吸速度が低下する、という二つの説がありました。しかし、CO2濃度を高くして光呼吸を阻害してもKok効果が見られること、O2濃度を下げて暗呼吸を阻害するとKok効果が見られなくなることから、暗呼吸速度が光強度が上がると阻害されることが原因であることがわかりました(Sharp et al. 1984)。
光照射時にどれだけ暗呼吸速度があるかは、低CO2濃度でCO2吸収速度を測定することによって得られます。光合成の二酸化炭素濃度依存性は次の項で説明していますが、CO2吸収速度はCO2濃度が下がると減少します。これは、光合成によるCO2吸収が減少することと、光呼吸によるCO2放出が高くなることの二つによります。40-50ppm前後でCO2吸収は0になります。この近辺でCO2濃度とCO2吸収速度の関係は直線です。しかし弱い光の下では、カルビンサイクルの酵素が不活性化されるため、光強度が変わると同じCO2濃度でも光合成速度が下がります。様々な光強度の下でCO2濃度とCO2吸収速度の関係を比べると、以下のグラフのようになります。
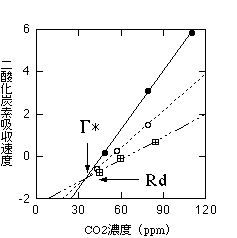
セイヨウキョウチクトウでの実測。RdはDay respiration。
両直線の交点となるCO2濃度では、光合成によるCO2吸収と光呼吸によるCO2放出がつりあっていると考えられます(ある程度光強度が強くなってしまうと、暗呼吸速度は光強度に依存しないと考えられているためです)。そこで、このCO2濃度を「暗呼吸速度を無視したときの光合成のCO2補償点」と呼びます。こちらでG*として表されているパラメータです。そして、このCO2濃度でのCO2放出速度が、光が当たっているときの暗呼吸速度(Day respirationと呼ばれています。日本語訳をみたことがありません)ということになります(Brooks & Farquhar 1985)。Day respirationは、暗呼吸速度のだいたい30-50%程度の速度であるようです(Villar et al. 1995)。Day respirationについてはいろいろ研究が進んでいますが、私はよく知りません。