光合成の生理生態学講座
はじめに (200509 24)
光合成を行う場 (200509 24)
チラコイド反応 (200509 24)
カルビンサイクル (200509 24)
糖合成 (200509 24)
光呼吸 (200509 26)
物質・エネルギー収支 (200509 26)
葉緑体の構造 (200509 26)
ここでは光合成の機作(メカニズム)を説明します。しかし「説明」と言ってもどこまで細かく説明するかは大きな問題です。この光合成の生理生態学講座は光合成のメカニズムを理解してもらうことが主目的ではありませんので、基礎的なところ、及び他の部分を理解してもらうために必要な部分を書くことにとどめます。
ここでは光合成系全体を俯瞰し、徐々に視点を細かくしていくことにします。光合成系は多くの構成要素から成り立っていますが、各構成要素についての細かい説明は「光合成構成要素」で説明することにします。
なお、ここで取り上げるのは高等植物のC3光合成です。また、光合成に付随する重要な代謝経路である光呼吸をここで取り上げます。
光合成の生化学的メカニズムについては、植物生化学(Heldt HW著、シュプリンガーフェアラーク東京)、植物生理学(Taiz L, Zeiger E編、培風館)、光合成事典(日本光合成研究会編、学会出版センター)をお勧めします。
光合成を行う場は、細胞内小器官の一種である葉緑体です。葉緑体は包膜で囲まれ、内部にチラコイド膜と呼ばれる膜構造を持っています。チラコイド膜と包膜の間の空間はストロマと呼ばれています(葉緑体の構造の説明はこちら)。
あとで詳しく書きますように、光合成反応は多くの構成要素がかかわる複雑な反応です。反応が起こる場所と役割を考えると、光合成反応は大きく二つの段階に分けることができます。
一つはチラコイド膜で起こる反応で、チラコイド反応と呼ばれます。ここでは光エネルギーを吸収し、化学エネルギーと還元力に変換されます。もう一つはストロマで起こる反応で、発見者の名をとってカルビン-ベンソンサイクル(回路)、あるいはカルビンサイクルと呼ばれます。このサイクルでは、チラコイド反応で生産された化学エネルギーと還元力を利用して、CO2が取り込まれトリオースリン酸(3つの炭素と1つのリン酸をもつ糖リン酸)が生産されます。このトリオースリン酸からスクロース(ショ糖)が合成されます。
上記のように、チラコイド反応の役割は、化学エネルギーと還元力の生産です。
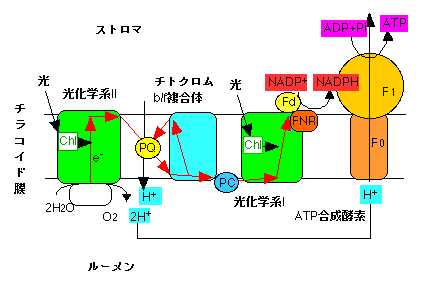
チラコイド反応は大きく電子伝達系とATP合成系に分けることができます。チラコイド膜上には4つの大きなタンパク質複合体があります。光化学系I複合体、光化学系II複合体、チトクロムb/f複合体、H+-ATP合成酵素複合体です。これらのうち光化学系I、光化学系II、チトクロムb/f複合体が電子伝達系の主要経路となります。
二つの光化学系にはクロロフィルという色素が結合しています。この色素が光を吸収し、励起されます。この励起エネルギーは、それぞれの光化学系の反応中心に送られます。光化学系の反応中心は励起エネルギーを受け取ると励起され、電子を放出します(光化学反応)。この放出された電子が電子伝達系内で伝達されるわけです。
光化学系IIの反応中心から電子が放出されると、電子を失った反応中心に電子が供給されます。この電子は水を分解することによって得られます。水2分子(2H2O)を分解すると、二つの電子(e-)、四つのプロトン(H+)、一つの酸素分子(O2)が発生します。光合成で発生する酸素はこれに由来します。
光化学系IIの反応中心から放出された電子は、光化学系II内部で伝達されたのち、プラストキノンという小分子に伝達されます。プラストキノンは電子を受け取ると同時にストロマ側にあるプロトンと結合し、チトクロムb/f複合体へと移動します。そしてチトクロムb/f複合体に電子を渡すと同時にルーメン内にプロトンを放出します。
電子はチトクロムb/f複合体からプラストシアニンを経て光化学系Iへ伝達されます。光化学系Iでは光化学系II同様に光化学反応が起こり、電子は光化学系Iからフェレドキシンという小さなタンパク質に伝達されます。この電子は最終的にNAPD+(ニコチンアミドジヌクレオチドリン酸)に渡され、NADPHが生産されます。このNADPHが還元力(電子供与体)としてカルビンサイクルで利用されます。以上が電子伝達系です。
上で述べたように、電子伝達においては、プラストキノンがプロトンをストロマからルーメンへ運搬します。また、ルーメンでは水分解の際にプロトンが放出されます。これらの二つの反応により、光合成がさかんに起こっているときにはルーメン内のプロトン濃度が上昇し、チラコイド膜の外側と内側でプロトンの濃度勾配が生じます。ルーメンのプロトンは、ATP合成酵素を通過してストロマに放出されます。この通過の際に、ATP合成酵素でATP(アデノシン三リン酸)が生産されます。ここでは、濃度勾配にそってプロトンが移動する際の運動エネルギーがATP合成に利用されていると考えて下さい。水力発電で水が上から下に移動するときのエネルギーが発電に利用されているようなものと考えていいと思います。ATPは生体内で「エネルギー通貨」として普遍的に利用されている分子で、カルビンサイクルでの化学エネルギーの供給源となります。
チラコイド反応は、かつて明反応と呼ばれていました。この呼び名は、光がないと反応が進まない、という意味で使われていましたが、現在ではあまり使われなくなっています。チラコイド反応の中でも本当に光が必要なのは光化学反応だけで、他の反応は光がなくても進むからです。
チラコイド反応のしくみ。PQはプラストキノン、PCはプラストシアニン、Fdはフェレドキシン、FNRはフェレドキシンNADP酸化還元酵素、Chlはクロロフィル。F0とF1はATP合成酵素の部分の名称。
カルビンサイクルの役割は、二酸化炭素を固定し、トリオースリン酸を生産することです。これにはエネルギーが必要で、チラコイド反応で生産されたNADPHとATPが利用されます。
カルビンサイクルは11の酵素からなる複雑な循環経路です。ここではポイントとなる反応についてのみ説明しましょう。
二酸化炭素の吸収は、リブロース二リン酸カルボキシラーゼ/オキシゲナーゼ(RuBPCase あるいはrubisco)と呼ばれる酵素が触媒するカルボキシル化反応(carboxylation)によって行われます。CO2はリブロース二リン酸(ribulose-1,5-bisphosphate, RuBP)という5つの炭素と2つのリン酸を持つ物質に結合し、2分子のホスホグリセリン酸(PGA)が生産されます。PGAは3つの炭素と1つのリン酸をもつ有機酸リン酸です。PGAから、ATPとNADPHを消費してトリオースリン酸(3つの炭素と1つのリン酸をもつ糖リン酸、TP)が生産されます。このTPの一部がスクロース合成に利用されます。残りのTPからはATPを消費してRuBPが再生され、再びカルボキシル化反応に使われることになります。
なお、デンプンが合成される際は、TPではなくフルクトース6-リン酸が利用されます。
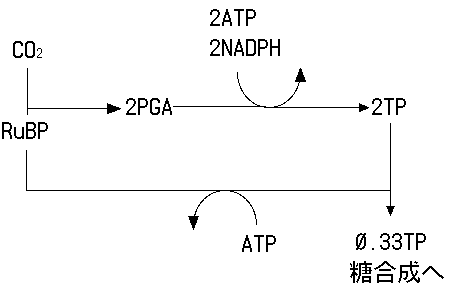 極めて単純化されたカルビンサイクル
極めて単純化されたカルビンサイクル
スクロース合成に利用されるトリオースリン酸(TP)は葉緑体外に運ばれます。その際葉緑体包膜にあるトリオースリン酸輸送体(リン酸トランスロケーター)を通過します。この輸送体はTP1分子を膜外に輸送する際に無機リン酸1分子を膜内にとりこみます。片方だけの輸送は行わないため、細胞質の無機リン酸濃度が低下するとTPを輸送できなくなります。
TPは細胞質でフルクトース二リン酸、フルクトースリン酸を経てスクロース(ショ糖)に合成されます。この際TPが持っていたリン酸が遊離し、TP輸送体を経由して葉緑体内に戻ります。
スクロースは細胞内外で利用されます。別の組織・器官へは師管を通じて輸送されます。この意味ではスクロースが光合成の最終産物であると考えていいのではないかと思います。
光合成ではデンプンも最終産物であると見なされています。デンプンは上記のようにカルビンサイクルのフルクトースリン酸から、葉緑体内で合成されます。デンプンは高分子(グルコースが連なったもの)であるため葉緑体包膜からそのままの形で出ることはありません。グルコースまで分解され、包膜のグルコース輸送体を通過して葉緑体外に出ることが多いようです。
RuBPCaseは、RuBPにCO2をくっつける反応(カルボキシル化)だけでなく、RuBPにO2をくっつける反応(酸素化反応、oxygenation)も触媒します。この反応の生成物は、1分子のPGAと1分子のグリコール酸リン酸(ホスホグリコール酸、以下PGlA)です。PGlAは2つの炭素と1つのリン酸をもつ物質です。PGlAからPGAを再生する系がグリコール酸回路です。PGlAは2つの炭素をもち、PGAは3つの炭素をもちます。1つのPGAを再生するためには2つのPGlAが必要で、途中で余った炭素がCO2として放出されます。CO2が放出される点が呼吸と似ているため光呼吸とも呼ばれるわけです。
しかし、通常の呼吸(暗呼吸、ミトコンドリア呼吸と呼ばれる)とは大きく違います。まず、代謝経路としては基本的に別のものです。次に、エネルギーの出入りが逆です。呼吸の役割は、糖を分解し、エネルギーをATPとして取り出すことです。一方、光呼吸ではPGlAからPGAを再生する際にエネルギー(ATP)と還元力(フェレドキシン<図中Fd>から電子を供給される)を消費します。
グリコール酸回路はカルビンサイクルよりもはるかに複雑な経路です。その中間代謝産物は葉緑体を出てミトコンドリア、ペルオキシソームと呼ばれる細胞内小器官で代謝されます。なお、CO2が放出される場所はミトコンドリアです。
光呼吸は、その名の通り光が当たっているときにしかおきません。グリコール酸経路が直接光を利用しているわけではないのですが、上記のようにグリコール酸回路を回すためには光のエネルギーが必要であることと、光がないとカルビンサイクルも回らないのでRuBPが生産されず、したがって酸素化反応も起きないからです。
光呼吸では、エネルギーが消費され、CO2も失われます。このため物質生産には全く貢献しません。光呼吸の役割はよくわかっていませんが、基本的には有害なグリコール酸リン酸を無事除去するためであると考えていいと思います。ではなぜRuBPCaseはわざわざ酸素化反応を触媒するのか、というと、「しかたなく」しているのではないかと思われます。カルボキシル化を触媒する構造を持つと酸素化の触媒が不可避である、ということですね。実際、様々な研究者が遺伝子改変によって酸素化を触媒しないRuBPCaseを作り出そうとしていますが、今のところうまくいった話は聞いていません。しかしなぜ酸素化反応が起こるのか、確かなところはわかっていません。
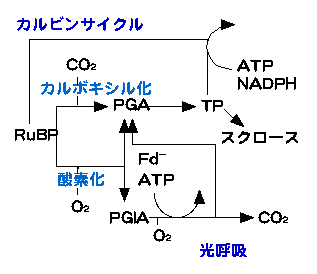 カルビンサイクルと光呼吸
カルビンサイクルと光呼吸
1分子のCO2の固定に伴ってどれだけの物質、エネルギーの出入りがあるかを考えてみましょう。
物質収支としては下式の通りで、この関係は基本的に一定、つまり1分子のCO2が吸収される際に、差し引き1分子の水が取り込まれ、1分子の酸素が発生します。
CO2 + 2H2O → (CH2O) + H2O + O2
もう少し細かく見てみましょう。
カルビンサイクルでは、CO2が1分子取り込まれると、合計3分子のATPと2分子のNADPHが消費されます。
電子伝達系では、1分子のNADP+を還元しNADPHを生産するために2つの電子が必要です。1つの電子を伝達するためには、二つの光化学系で光化学反応が起こる必要があります。したがって少なくとも2つの光量子が必要になります。つまり、NADPHの供給がカルビンサイクルを規定しているとすると、1分子のCO2を固定するために最低8光量子が必要であることになります。
1分子のATPを合成するためにどれだけの光量子が必要なのかは、まだ正確にはわかっていません。まず、1光量子あたりでどれだけのプロトン輸送ができるのかがわかっていません。ルーメンにおけるプロトンの増加には、水分解とプラストキノンによる輸送がかかわっています。水分解のほうは難しくありません。酸素が1分子発生する際には2分子の水が分解され、4つのプロトンと4つの電子が生じます。上記のように1つの電子を伝達するためには最低2つの光量子が必要で、1プロトンの発生に2光量子(以上)が必要であることがわかります。一方、プラストキノンの輸送ですが、電子伝達系には上で説明した直列的〔非循環型、開放型とも言う)電子伝達の他に、循環的電子伝達というのが何種類かあります。ここでは詳しく説明しませんが、電子伝達の様式によってプラストキノンによるプロトン輸送の効率が変わると理解して下さい。直列的電子伝達が最も効率よくはたらいているときには、おそらく1電子(つまり2光量子)あたり2つのプロトンが輸送されます。これを総合すると、2光量子で最大3プロトンをルーメンにため込むことができます。次にATP合成とプロトンの関係ですが、1分子のATPを生産するために必要なプロトン輸送量がよくわかりません。昔は3プロトンから1ATPが合成されると考えられていたようですが、最近4プロトンから1ATPが合成される、という報告があったようです(Haraux and De Kouchkovsky 1998)。どれが正しいのかは私にはよくわかりませんが。ATPの供給がカルビンサイクルを規定しているとすると、これらの合計から、1分子のCO2を固定するために8光量子が必要である、ということになります。
注意してほしいのですが、3ATP合成に8光量子、2NADPH合成に8光量子別々に必要なわけではありません。8光量子で3ATPと2NADPHが合成され、1CO2がめでたく合成される、ということです。
では、光合成をエネルギー効率という点から見てみましょう。光量子は波長によって持っているエネルギーが違います。波長が短い(紫とか青とか)場合は1光量子あたりのエネルギーが多く、長い(赤)場合は小さくなります。光化学反応は波長の長さには依存せず、青でも赤でも1つの光量子で光化学反応を起こすことができます。赤い光(例として波長680nm)の場合、1光量子あたり175kJのエネルギーを持ちます。これに対し、ATPの合成には29kJ、NADPHの合成には217kJが必要です。1CO2を固定するのに必要な3ATPと2NADPを生産するためには521kJあれば充分ですが、実際に投入されているエネルギーは8光量子で1400kJの光エネルギーが利用されていることになります。つまりチラコイド反応のエネルギー効率は最大でも37%程度ということになります。一方、カルビンサイクルで1分子のCO2を糖に変換する際には約480kJのエネルギーが必要です。投入されているのは521kJですから、カルビンサイクルのエネルギー変換効率は90%以上ということになります。これらから、光合成のエネルギー変換効率は最大でも34%前後ということになります。
しかしこの値はあくまで理論値であり、現実にこのような効率でCO2が固定されることはありません。以下にいくつか効率を下げる要因を挙げてみましょう。
・波長
上の例は最もエネルギーが低い赤色光の話ですが、波長が低い光は1光量子あたりのエネルギーは多くなります。波長が短い光がクロロフィルに吸収された場合、余分なエネルギーは熱となって消えていきます。
・光吸収→光化学反応
光化学系IIでは、光強度が非常に弱い場合、クロロフィルに吸収されたエネルギーの約80%が光化学反応に使われ、残りの20%は熱や蛍光として消えていきます(赤色光の場合)。また、光強度が強くなると、効率は大きく落ちます。光化学反応の下流(電子伝達やカルビンサイクル)の反応速度が追いつかないためです(このような状態を光飽和といいます)。この場合は、吸収されたエネルギーは使われず、熱として放散されていきます。
・光呼吸
通常のCO2濃度(370ppm)では、RuBPのカルボキシル化と酸素化の両方が起きます。上記の通り、光呼吸ではせっかく固定したCO2が失われ、エネルギーが消費されます。カルボキシル化と酸素化の割合は条件によって異なりますが、おおざっぱに言って光呼吸によって30-40%のエネルギーが無駄になっています。
この他、チラコイド膜からのプロトンのリーク、循環的電子伝達などが考えられますが、ここではふれないでおきます。
1光量子あたりどれだけのCO2が吸収できるかを量子収率と呼びます。最大可能量子収率は1/8=0.125であることになりますが、現実の植物でどうなるかは、「光と光合成」で述べることにしましょう。
光合成は葉緑体で行われます。うまく操作すれば細胞から単離した葉緑体の光合成を測定することもできます(私はしたことないんですが)。ということで、葉緑体は光合成を行う最小単位ということになります。
ここでは、葉緑体の形態を主に紹介しましょう。単に形がどうなっているかだけではなく、それが機能とどう結びついているか、まで書きたいと思います。
葉緑体の構造の模式図は下の通りです。
通常、断面図が描かれることが多いため、葉緑体は棒状だと思われがちですが、高等植物の場合、葉緑体は扁平であることが多いようです。
葉緑体は包膜で包まれています。包膜は二重の生体膜からなります(ただし、ぴったりくっついているため二重に見えるわけではないようです)。包膜上には様々なイオンなどのトランスポータがあります。
葉緑体内部にはチラコイド膜と呼ばれる膜構造があります。チラコイド膜は袋状に閉じていて、内部と外部はそれぞれ隔離されています。チラコイド膜の外側をストロマ、内側をルーメンと呼んでいます(ちなみに、「ストロマ」は葉緑体特有の名称ですが、「ルーメン」は一般名詞です)。ストロマにはカルビンサイクルがあることは上に述べた通りです。
チラコイド膜は層状に重なったグラナ(grana, grana stacking)とよばれる構造を作ります。このグラナ構造は座布団が積み重なっているように見えますが、座布団と違って、一つのグラナのチラコイド膜はつながっているらしいです。グラナの積み重なりの維持にはマグネシウムイオンが必要で、チラコイド膜をマグネシウムイオンがないバッファに入れてやるとグラナは解消されます。グラナ部分のチラコイド膜をグラナチラコイド、それ以外の部分のチラコイド膜をストロマチラコイドと言います。
チラコイド膜には4つの大きなタンパク質複合体が存在することは上で述べました(光化学系I・II・チトクロムb/f・ATPase)。これらの複合体の膜状の分布は均一ではありません。光化学系IIのほとんど(LHCIIを多く持っている光化学系IIで、PSIIaという)は、グラナ中のチラコイド膜とチラコイド膜が接している部分(appressed regionという)に分布します。光化学系IIの一部(LHCIIが少ない。PSIIbという)・光化学系I・ATPaseはストロマと接している側(non-appressed region)にのみ分布します。チトクロムb/f複合体のみはどちらの領域にも分布しています。
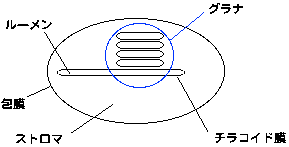 葉緑体の模式図
葉緑体の模式図