岝崌惉偺惗棟惗懺妛島嵗
丂
栚師
丂
偼偠傔偵丂乮200709 15乯
梩孮岝崌惉懍搙偺寁嶼尨棟乮200709 15乯
梩孮撪偺屄梩岝崌惉摿惈乮200710 15乯
丂媧岝學悢偲梩柺愊巜悢乮200711 27乯
丂拏慺暘攝乮200712 25丒200801 06乯
丂拏慺棙梡傪峫椂偟偨嵟揔梩柺愊巜悢乮2009 0115乯 New
丂摦揑側梩孮岝崌惉儌僨儖
丂
丂
丂梩偭傁堦枃偟偐傕偨側偄怉暔丄偲偄偆偺傕偄傞偵偼偄傑偡偑丄懡偔偺怉暔偼偨偔偝傫偺梩傪帩偪傑偡丅傑偨丄偩偄偨偄偵偍偄偰怉暔偼孮棊傪宍惉偟偰惗妶偟偰偍傝丄偁傞堦掕柺愊偺拞偵偼偨偔偝傫偺梩偑偁傞偙偲偑傢偐傝傑偡丅惗懺宯偺暔幙弞娐傪峫偊傞偲偒丄怉暔偺戝偒側栶妱偺堦偮偼岝崌惉傪偟偰丄扽慺傪庢傝崬傓偙偲偱偡丅岝崌惉傪偡傞扨埵偲偟偰孮棊傪尒傞偲丄孮棊偲偄偆偺偼梩偺廤傑傝偱偡丅屄乆偺梩偺岝崌惉偺憤榓偑梩孮岝崌惉乮孮棊岝崌惉乯偱偡偑丄偙偪傜偱愢柧偟偨傛偆偵丄屄乆偺梩偺旝娐嫬丄摿偵岝娐嫬偼戝偒偔堎側傝丄偦傟偵埶懚偟偰岝崌惉検偼堎側傝傑偡丅杮崁偱偼丄梩孮岝崌惉傪梩偺岝崌惉偺憤榓偲峫偊偨偲偒偵崻掙傪巟攝偡傞榑棟傪峫嶡偟傑偡丅擄偟偄尵偄曽偱偡偑丄梩孮岝崌惉偵塭嬁傪梌偊傞梫場偵偮偄偰抦偭偰傕傜偍偆丄偲偄偆偙偲偱偡丅
丂側偍丄乽梩孮岝崌惉乿偼canopy photosynthesis偺栿偲偟偰弌偟傑偟偨丅姷廗偲偟偰偼乽孮棊岝崌惉乿偺傎偆偑側偠傫偱偄傑偡偑丄偙偙偱愢柧偡傞儌僨儖偼梩偺傒傪埖偄丄宻丒崻側偳偼柍帇偟偰偄傞偺偱丄乽梩孮岝崌惉乿偺傎偆偑懨摉偲偄偆偙偲偱偡丅
丂杮崁傪撉傓慜偵丄偙偪傜傪撉傒丄梩孮撪偺岝娐嫬偺晄嬒堦惈偵偮偄偰棟夝偟偰偔偩偝偄丅
丂
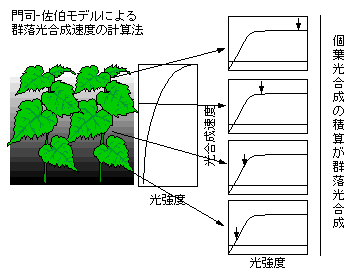
丂梩孮傪岝崌惉偺扨埵偲偟偰弶傔偰峫偊偨偺偼儃僀僙儞-僀僃儞僙儞偱偁傠偆偲巚傢傟傑偡乮Boysen Jensen 1932. 偪側傒偵乽Boysen Jensen乿偑惄丅乯丅斵偼僔儘僈儔僔側偳偺彫偝側敨怉偊孮棊傪嶌傝丄偦偺岝崌惉懍搙傪應掕偟傑偟偨丅偝傜偵屄梩偺岝崌惉懍搙偲偺斾妑傪峴偄丄梩柺愊偁偨傝偺屄梩岝崌惉懍搙偵斾傋梩柺愊偁偨傝偺梩孮岝崌惉懍搙偑掅偄偙偲傪帵偟丄梩孮岝崌惉偑屄梩岝崌惉偺扨弮側榓偱偼側偄偙偲傪巜揈偟傑偟偨丅偙傟偼丄斵帺恎偑峫嶡偟偨傛偆偵丄梩孮撪偺岝娐嫬偺晄嬒堦惈側偳丄屄梩岝崌惉傪應掕偡傞偲偒偵偼柍帇偝傟偰偄偨梫場偑梩孮岝崌惉懍搙偵戝偒側塭嬁傪梌偊傞偨傔偱偡丅
丂梩孮岝崌惉懍搙偵偳偺傛偆側梫場偑塭嬁偡傞偺偐丄偵偮偄偰偺棟榑揑側榞慻傒偼Monsi and Saeki (1953) 偵傛偭偰幚尡揑偐偮棟榑揑偵柧傜偐偵偝傟傑偟偨丅斵傜偼偙偪傜偱愢柧偟偨傛偆側梩孮撪偺岝娐嫬暘晍傪幚應丒棟榑偺椉柺偐傜柧傜偐偵偟丄偝傜偵丄屄梩偺岝崌惉摿惈偲岝娐嫬暘晍傪岻傒偵慻傒崌傢偣傞偙偲偵傛傝丄梩孮慡懱偺岝崌惉懍搙傪儌僨儖壔偡傞偙偲偵惉岟偟傑偟偨丅
丂Monsi and Saeki (1953) 偺梩孮岝崌惉儌僨儖乮孮棊岝崌惉儌僨儖乯偺尨棟傪娙扨偵徯夘偡傞偲埲壓偺捠傝偵側傝傑偡丅傑偢丄梩孮撪偵偼岝娐嫬偺岡攝偑偁傝傑偡丅偁傞憌n偺梩偑庴偗傞梩柺愊偁偨傝偺岝嫮搙Iincn傪Iincn=K Io exp(-K Fn)偲偟偰寁嶼偟傑偡乮徻偟偔偼偙偪傜乯丅偦偟偰奺梩偺岝皩鯈噽瑡葠鼈饓紥鑲祩軅穪iMonsi and Saeki 儌僨儖偱偼捈妏憃嬋慄偲捈慄傪慻傒崌傢偣偨傕偺偑巊傢傟偨乯丅Monsi and Saeki 儌僨儖偱偼慡偰偺梩偑摨偠岝皩鯈噽瑡葠鼈饚潅聜茐紥鑲祦A奺憌偺岝嫮搙偲岝-岝崌惉嬋慄偐傜丄奺憌偺梩偺岝崌惉懍搙傪寁嶼偟傑偟偨丅偦偟偰丄慡偰偺梩偺岝崌惉懍搙偺愊暘偑梩孮岝崌惉懍搙乮孮棊岝崌惉懍搙乯偲偟偰寁嶼偝傟偨傢偗偱偡丅
丂梩孮岝崌惉懍搙偼堦斒偵搚抧柺愊偁偨傝偱昞偝傟傑偡丅偙偺儌僨儖偱偼搚忞傗崻丒宻偺屇媧偼柍帇偝傟偰偍傝丄傕偭傁傜梩偺扽慺廂巟偺傒偑寁嶼偝傟傑偡丅
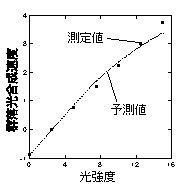
丂Monsi and Saeki (1953) 偺榑暥偑彂偐傟偨摉帪偼愴憟捈屻偱偁傞偙偲傕偁傝岝崌惉懍搙傪應掕偡傞僔僗僥儉偑側偔丄斵傜偼儃僀僙儞-僀僃儞僙儞偑應掕偟偨僔儘僈儔僔側偳偺屄梩岝崌惉懍搙傪梡偄偰梩孮岝崌惉懍搙傪寁嶼偟丄幚應偝傟偨梩孮岝崌惉懍搙偲斾妑偡傞偲偄偆偙偲傪偟偰偄傑偡丅壓偺恾偼Monsi and Saeki (1953) 偵嵹偭偰偄傞幚應丒梊應偝傟偨岝崌惉懍搙偺斾妑偱偡丅梊應偼旕忢偵偆傑偔幚應抣偲偁偭偰偍傝丄儌僨儖偑懨摉偱偁傞偙偲傪帵嵈偟偰偄傑偡丅
丂偁偲偱弎傋傑偡傛偆偵丄梩孮岝崌惉儌僨儖偵偼偙偺偁偲條乆側夵椙偑巤偝傟偰偄傑偡丅偟偐偟婎杮亅梩孮撪偺岝娐嫬偲屄梩偺岝-岝崌惉嬋慄偐傜屄梩偺岝崌惉偑寁嶼偝傟丄偦偺榓偲偟偰梩孮岝崌惉懍搙偑寁嶼偝傟傞-偼尰嵼傕曄傢偭偰偄傑偣傫丅
丂
丂Monsi and Saeki (1953) 偺儌僨儖偱偼丄慡偰偺梩偑摨偠岝-岝崌惉嬋慄傪帩偮偙偲偑壖掕偝傟偰偄傑偟偨丅偙偺壖掕偑惓偟偔側偄丄偮傑傝梩偵傛偭偰岝崌惉摿惈偑堎側傞偙偲偼偡偱偵斵傜帺恎偑抦偭偰偍傝丄椺偊偽Saeki (1959) 偼働僀僩僂偺孮棊撪偱忋晹偺庒偄梩偺岝崌惉擻椡乮岝朞榓壓偺岝崌惉懍搙乯傗屇媧懍搙偑壓晹偺擭庢偭偨梩偵斾傋崅偄偙偲傪曬崘偟偰偄傑偡丅斵傜偑丄偙偺傛偆側惓偟偔側偄偲傢偐偭偰偄傞壖掕傪梡偄偨棟桼偼偍偦傜偔擇偮偁傝傑偡丅堦偮偼寁嶼擻椡偺尷奅偱偡丅摉帪僐儞僺儏乕僞側偳偲偄偆曋棙側婡夿偼傑偩側偔丄斵傜偼寁嶼広傗庤夞偟寁嶼婡側偳偱暋嶨側寁嶼傪偡傞昁梫偑偁傝傑偟偨丅寁嶼幃偼偱偒傞尷傝夝愅揑偵夝偒丄夝偔偺偑晄壜擻側晹暘傪寁嶼婡偱曗偆偙偲傪偟偰偄偨偺偱偼側偄偐偲巚傢傟傑偡丅偦偺偨傔寁嶼傪扨弮壔偝偣傞偙偲偼戝帠側栤戣偱偁偭偨偲憐憸偝傟傑偡丅傕偆堦偮偼丄偙偺傛偆側壖掕偱傕梩孮岝崌惉懍搙傪偐側傝偺惛搙偱悇掕偡傞偙偲偑壜擻偱偁偭偨偐傜偩偲巚傢傟傑偡丅梩孮撪偱偼壓晹偺梩偺岝娐嫬偼埆偄偨傔岝崌惉偼岝棩懍偱偁傝丄岝崌惉擻椡偺嵎偑尰傟偵偔偄偙偲丄傑偨丄忋晹偺梩偲壓晹偺梩傪斾傋傞偲丄庛岝壓偱偺岝崌惉懍搙偵偼偁傑傝戝偒側嵎偑側偄偨傔丄忋晹偺梩偲摨偠惈幙傪壖掕偟偰傕岆嵎偼偁傑傝戝偒偔側傜側偄偺偱偡丅Saeki (1959) 偼丄梩孮岝崌惉懍搙傪悇掕偡傞偨傔偵偼丄梩孮嵟忋晹偺梩偺岝崌惉擻椡偲梩孮偺梩偺屇媧懍搙偺暯嬒抣偑傢偐傟偽廫暘偱偁傞偲弎傋偰偄傑偡丅偟偐偟幚嵺偵偼梩孮撪偺屄梩偺岝崌惉摿惈偵偼戝偒側堘偄偑偁傝丄屻弎偡傞傛偆偵偦傟偼怉暔偺帒尮棙梡偲嫮偔娭楢偑偁傝傑偡丅
丂偙偪傜偱弎傋偨傛偆偵丄梩孮撪偵偼岝娐嫬偵戝偒側偽傜偮偒偑偁傝傑偡丅懡偔偺応崌丄屄梩偺岝崌惉摿惈偼岝娐嫬偵埶懚偟偰堎側傝丄椺偊偽忋晹偺梩傎偳岝崌惉擻椡丒屇媧懍搙丒拏慺娷検乮梩柺愊偁偨傝乯偑崅偄側偳丄偄傢備傞梲梩偲堿梩偺堘偄偑尒傜傟傑偡乮Hirose and Werger 1987a,b, 1988乯丅梲梩偲堿梩偵偮偄偰偼徻嵶傪偙偪傜偵彂偒傑偟偨偺偱偛棗偔偩偝偄丅
丂偨偩偟丄梩孮撪偺忋晹-壓晹偺岝崌惉摿惈偺堘偄偼丄慡偰梲梩-堿梩偺堘偄偲摨偠偱偁傞傢偗偱偼偁傝傑偣傫丅憪杮孮棊偱偼丄懡偔偺怉暔偑忋晹偵梩傪揥奐偟傑偡丅偁傞帪婜偵庒偔嵟忋晹偵偁偭偨梩偼丄怴偟偄梩偑忋晹偵揥奐偝傟傞偨傃偵旐堿偵傛偭偰岝娐嫬偑埆偔側偭偰偄偒傑偡丅偙偺傛偆側働乕僗偱偼丄壓晹偺梩偲偄偊偳梩偑嶌傜傟偨偲偒偼岝娐嫬偑椙偐偭偨偨傔丄梩偼嫮岝偱崅偄岝崌惉懍搙傪幚尰偱偒傞傛偆岤偔側偭偰偄傑偡丅梩偺宍懺偼堦搙偱偒偰偟傑偆偲偁傑傝曄傢傜側偄偨傔丄壓晹偺梩偱傕忋晹偺梩偲摨條岤偄偙偲偵側傝傑偡乮Hirose et al. 1988乯丅堦曽丄怷椦偱偼怉暔偺惉挿偼偦傟傎偳懍偔側偔丄梩偺岝娐嫬偼偁傑傝曄壔偟傑偣傫丅偙偺偨傔丄忋晹偺梩偼岤偔壓晹偺梩偼敄偄偲偄偆梲梩-堿梩偺堘偄偑尒傜傟傑偡乮Ellsworth and Reich 1993)丅怷椦庽栘偺梩孮撪偺屄梩偺惈幙偵偮偄偰偼懡偔偺尋媶偑偁傝丄摿偵僄僗僩僯傾偺Ulo Niinemets偑惛椡揑偵尋媶傪峴偄傑偟偨丅懡偡偓偰偙偙偱偼徯夘偟偒傟側偄偺偱嵟嬤弌偨儗價儏乕傪帵偟偰偍偒傑偡乮Niinemets and Valladares 2004, Niinemets 2007乯丅
丂
丂
丂梩孮岝崌惉懍搙偵偼條乆側梫場偑塭嬁偟傑偡丅Monsi-Saeki 儌僨儖傪巊偭偨弶婜偺尋媶偱偼丄庡偲偟偰梩孮峔憿偵拝栚偟丄媧岝學悢偲梩柺愊巜悢偑梩孮岝崌惉懍搙偵梌偊傞塭嬁偑尋媶偝傟傑偟偨丅偙偺夁掱偱惗傑傟偨奣擮偑嵟揔梩柺愊巜悢偱偡丅梩偺岝崌惉懍搙偼丄庛偄岝傎偳掅偔側傝丄偁傞岝嫮搙乮岝曗彏揰乯傛傝傕掅偄偲屇媧懍搙偑岝崌惉懍搙傪忋夞傞傛偆偵側傝傑偡丅Beer偺朄懃偵廬偊偽丄孮棊偺嵟壓晹偺梩偑庴偗傞岝嫮搙偼孮棊偑帩偮梩柺愊巜悢偑懡偄傎偳掅偔側傝傑偡丅梩偑庴偗傞岝嫮搙偑僾儔僗偺CO2媧廂傪堐帩偱偒側偄偔傜偄掅偄偲丄偦偺梩偼岝崌惉惗嶻偵偼栶偵棫偪傑偣傫丅Monsi-Saeki儌僨儖偵偟偨偑偊偽丄嵟壓憌偺梩偑偓傝偓傝僾儔僗偺CO2媧廂傪堐帩偱偒傞掱搙偵梩柺愊巜悢傪帩偰偽丄偦偺孮棊偺岝崌惉検偑嵟戝偵側傝丄偦傟埲忋梩傪帩偮偲丄偐偊偭偰孮棊岝崌惉偼掅偔側傝傑偡丅孮棊岝崌惉傪嵟戝偵偡傞梩柺愊傪乽嵟揔梩柺愊巜悢乿偲屇傃傑偡丅
丂嵟揔梩柺愊巜悢偼梩偺岝崌惉摿惈乮岝曗彏揰-岝崌惉懍搙偑0偵側傞岝嫮搙乯偲媧岝學悢偵埶懚偟傑偡丅慡偰偺梩偺岝崌惉摿惈傪堦掕偵偟偨応崌丄梩柺愊巜悢偲孮棊岝崌惉偺娭學偼埲壓偺傛偆偵側傝傑偡丅
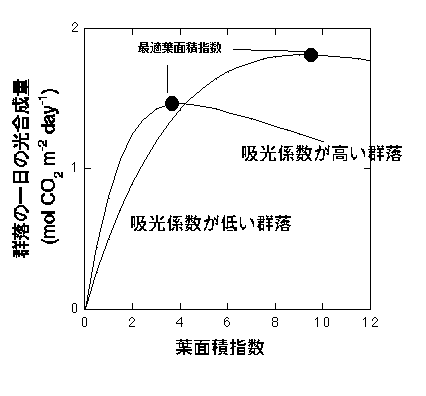
梩孮岝崌惉懍搙偲梫柺愊巜悢偺娭學丅愄偁傞杮偺復傪彂偔偨傔偵寁嶼偟偨傕偺偺挿偔側偭偰偟傑偆偨傔儃僣偵側偭偨寁嶼丅偙偪傜偵帵偟偨僰儅僈儎偲僆僆僆僫儌儈孮棊偺僨乕僞傪傕偲偵寁嶼偟偨傕偺偱偡偑丄徻嵶偼朰傟傑偟偨丅
恾偵偁傞傛偆偵丄嵟揔梩柺愊巜悢偼媧岝學悢偑崅偔側傞偲掅偔側傝傑偡丅偙傟偼媧岝學悢偑掅偄偲梩孮偺壓晹傊撏偔岝偑彮側偔側傝丄梩柺愊巜悢偑掅偔偲傕岝曗彏揰傪壓夞偭偰偟傑偆偨傔偱偡丅傑偨丄嵟揔梩柺愊巜悢偑幚尰偝傟偰偄傞応崌丄梩孮岝崌惉懍搙偼媧岝學悢偑崅偄孮棊傎偳崅偔側傝傑偡丅嵟揔梩柺愊悢偑幚尰偝傟偰偄傞忬懺偱偼丄梩孮偑媧廂偡傞岝偺検偼堦掕偱偡乮嵟壓晹偺梩偑媧廂偡傞岝偑岝曗彏揰乯丅偮傑傝丄摨偠岝傪媧廂偟偰偄傞偺偵媧岝學悢偑崅偄梩孮偺傎偆偑岝崌惉懍搙偑崅偄偺偱偡丅偙傟偑側偤偐偲偄偆偺偼彮偟擄偟偄偱偡偑丄棟孅偼埲壓偺捠傝偱偡丅傑偢丄媧岝學悢偑掅偄梩孮偵斾傋丄媧岝學悢偑崅偄梩孮偱偼丄嵟忋晹偺梩偑媧廂偡傞岝偑嫮偔側傝傑偡乮偙偪傜嶲徠乯丅偟偐偟丄偛懚偠偺捠傝岝崌惉懍搙偼偁傑傝岝偑嫮偄偲朞榓偟丄岝嫮搙偵埶懚偟側偔側傝傑偡丅偮傑傝嵟忋晹偺梩偺岝崌惉検偼丄傕偲傕偲岝偑嫮偄偨傔梩偺妏搙偵戝偒側塭嬁傪庴偗傑偣傫丅堦曽丄壓晹偺梩偺岝崌惉懍搙偼丄梩偺妏搙偵戝偒側塭嬁傪庴偗傑偡丅媧岝學悢偑崅偄孮棊偱偼壓晹偵傕傟傞岝偑彮側偄偨傔岝崌惉検偑彮側偄偺偵懳偟丄媧岝學悢偑掅偄孮棊偱偼岝崌惉検偑懡偔側傝傑偡丅偙偺嵎偑梩孮岝崌惉懍搙偵傂傃偒丄媧岝學悢偑掅偄孮棊偺傎偆偑岝崌惉検偑懡偔側傝傑偡丅梫偡傞偵丄媧岝學悢偑崅偄偲孮棊岝崌惉偺嵟戝抣偑掅偄偺偼丄嵟忋晹偺梩偼嫮偄岝傪媧廂偱偒傞傕偺偺丄慡偰偺岝傪棙梡偱偒傞傢偗偱偼側偄乮岝崌惉偑摢懪偪偵側偭偰偄傞乯偺偱丄摨偠岝検傪媧廂偟偰傕丄柍懯偑懡偄偐傜偱偡丅
丂嵟揔梩柺愊巜悢偺懚嵼偵偮偄偰偼丄條乆側媍榑偑偁偭偨傛偆偱偡丅幚尡揑偵偼丄媧岝學悢偺堎側傞庬傪巊偭偰條乆側枾搙偺孮棊傪嶌偭偨傝乮Watson & Wats 1959乯丄僀僱偺梩偵廳傝傪偮偗偰梩偺妏搙傪挷愡偟丄媧岝學悢偺堎側傞孮棊傪恖堊揑偵嶌偭偨傝乮Tanaka 1972; Monsi et al. 1973乯偟偰丄忋婰偺傛偆側孮棊岝崌惉-梩柺愊巜悢娭學傪幚應偟偨椺偑偁傝傑偡丅堦曽丄棟榑揑側柺偐傜嵟揔梩柺愊巜悢傪斲掕偟偨媍榑傕偁傝傑偟偨丅McCree偼丄孮棊壓晹偺梩傎偳岝崌惉偺岝曗彏揰偑掅偄偙偲傪巜揈偟傑偟偨丅Monsi-Saeki儌僨儖偱偼丄孮棊偺慡偰偺梩偑摨偠岝崌惉摿惈傪帩偮偲壖掕偟偰偄傑偡偑丄尰幚偼忋偵彂偄偨偲偍傝丄孮棊壓晹偺梩傎偳屇媧懍搙偑掅偄偲偄偆摿惈偑偁傝傑偡丅嵟揔梩柺愊巜悢偑側偔側傞偐偳偆偐偼暿偲偟偰丄屇媧懍搙偺曄壔傪擖傟傞偲嵟揔梩柺愊巜悢偺抣偑戝偒偔曄傢傞偺偼妋偐偱偡丅偙偙傑偱偺媍榑偼60-70擭戙偵偐偗偰峴傢傟偨傕偺偱偡乮崟娾愭惗偺嫵壢彂偵徻偟偄偄偒偝偮偑嵹偭偰偄傑偡乯丅偙傟傜偺栤戣偼傎偲傫偳朰傟傜傟偰偄偨偲偄偭偰偄偄偲巚傢傟傑偡偑丄80擭戙偵側偭偰丄怴偨側栤戣偑弌偰偒傑偟偨丅偦傟偑丄孮棊偑帩偮拏慺偺栤戣偱偡丅
丂
忋偱弎傋偨傛偆偵丄岝崌惉摿惈偵偼梩孮撪偱堘偄偑偁傝傑偡偑丄Monsi&Saeki儌僨儖埲崀丄懡偔偺梩孮岝崌惉儌僨儖偱偼柍帇偝傟偰偒傑偟偨乮椺偊偽Johnson and Thornley 1984乯丅
丂堦曽丄梩孮岝崌惉偺暘栰偲偼暿偵丄岝崌惉摿惈偺庬娫嵎傗娐嫬墳摎傪恑壔惗懺妛揑乮暔幙宱嵪揑乯娤揰偐傜棟夝偟傛偆偲偄偆帋傒偑70擭戙屻敿偐傜巒傑傝傑偟偨丅偙傟偑偙偪傜偱徯夘偟偨僐僗僩亅儀僱僼傿僢僩夝愅偱偡乮Mooney and Gulmon 1979, 1982乯丅偙偺峫偊曽偐傜丄岝崌惉傪棙摼丄拏慺傪僐僗僩偲傒側偡峫偊曽偑敪揥偟傑偡丅偙偺壖愢偼椺偊偽Gulmon & Chu (1981)傗DeJong & Doyle (1985)側偳偑岝娐嫬偲拏慺娷検偺憡娭傪帵偡偙偲偵傛傝巟帩偝傟偰偄偒傑偟偨丅偝傜偵丄Mooney偺僌儖乕僾偺堦堳偱偁傞Field (1983) 偼丄屄懱偺岝崌惉傪嵟戝壔偡傞拏慺暘攝偵偮偄偰峫嶡偟傑偟偨丅岝崌惉偲拏慺娷検偺娫偵娭悢娭學偑偁傞偲偡傞偲丄棟榑揑偵偼丄屄懱撪偺拏慺傪堦掕偲偟偨偲偒丄屄懱偺岝崌惉検偼丄屄懱撪偺慡偰偺梩偑埲壓偺幃傪枮偨偡傛偆偵拏慺傪暘攝偡傟偽嵟戝偵側傝傑偡丅
丂
兩= 佪A/佪N=堦掕
丂
偙偙偱丄A偲N偼屄梩偺岝崌惉検偲拏慺娷検偱偡丅儔儉僟偼儔僌儔儞僕僃偺忔悢偱丄掕悢偱偡丅偙傟傪恾傪巊偭偰愢柧偟傑偟傚偆丅
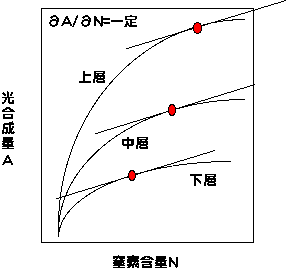
榖傪娙扨偵偡傞偨傔偵丄梩孮偑俁憌偐傜側傞偲峫偊傑偟傚偆丅偦偟偰丄奺憌偵偍偗傞拏慺娷検偲岝崌惉検偺娭學傪悇掕偟傑偡丅偙偪傜偱弎傋傑偟偨傛偆偵丄拏慺娷検偑憹偊傞偲岝崌惉擻椡偑忋偑傝傑偡丅偙偺偨傔拏慺偑憹偊傞偲岝崌惉検偑忋偑傝傑偡丅拲堄偟側偗傟偽偄偗側偄偺偼丄憌偵傛偭偰岝忦審偑堘偆偨傔丄摨偠拏慺娷検偱傕憌偵傛偭偰岝崌惉検偑堎側傞偙偲偱偡丅壓憌偺梩偼庛偄岝偟偐庴偗傑偣傫偑丄庛岝壓偺岝崌惉懍搙偼拏慺傪憹傗偟偰傕偦傟傎偳曄壔偑側偔丄摨偠拏慺娷検偱偼丄拏慺検憹壛偵懳偡傞岝崌惉検偺憹壛乮佪A/佪N乯偑彫偝偔側傝傑偡丅偟偐偟壓憌偺梩偱傕拏慺娷検偑掅偄偲佪A/佪N偑忋偑傝傑偡丅偮傑傝兩傪堦掕偵偡傞偲偄偆偙偲偼丄忋憌偵懡偔丄壓憌偵彮側偔拏慺傪暘攝偡傞偙偲偵側傝傑偡丅恾拞偺愒娵偑兩傪堦掕偵偡傞揰偱偡丅奺梩偵偙偺傛偆偵拏慺傪暘攝偡傞偲丄梩孮岝崌惉懍搙偑嵟戝偵側傝傑偡丅拲堄偟側偗傟偽偄偗側偄偺偼丄嵟揔暘攝偼梩孮偑傕偭偰偄傞拏慺検偵埶懚偡傞偲偄偆偙偲偱偡丅梩孮偑傛傝懡偔偺拏慺傪帩偭偰偄傟偽丄愒娵偼慡懱揑偵塃偵偢傟傑偡乮兩偑彫偝偔側傞乯丅
丂忋偺榖偼偐側傝崅摍側悢妛偱丄巹偵傕偒偪傫偲棟夝偱偒偰偄傞傢偗偱偼偁傝傑偣傫丅偦偙偱丄捈娤揑偵棟夝偡傞偨傔偵埲壓偺巚峫幚尡傪偟傑偟傚偆丅
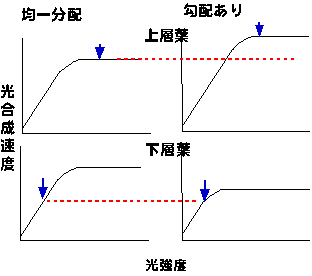
崱搙偼傕偭偲扨弮偵丄擇憌偐傜側傞梩孮傪峫偊傑偡丅嵍懁偺梩孮偱偼忋壓憌偲傕摍検偺拏慺傪傕偮偲偟傑偡乮嬒堦暘攝乯丅堦曽塃懁偺梩孮偱偼拏慺傪忋憌偵懡偔壓憌偵彮側偔傕偪傑偡乮岡攝偁傝乯丅偙偺偲偒梩孮慡懱偺岝崌惉検偼偳偆側傞偱偟傚偆偐丅徻偟偔偼偙偙偱偼愢柧偟傑偣傫偑丄庛岝壓偺岝崌惉懍搙偼拏慺娷検偑曄傢偭偰傕偁傑傝曄傢傝傑偣傫丅偙偺偨傔丄嵍塃偳偪傜偺梩孮偲傕壓憌偺梩偺岝崌惉検偼偦傟傎偳曄傢傝傑偣傫丅堦曽丄忋憌梩偼嫮偄岝傪庴偗傑偡丅嫮岝壓偺岝崌惉懍搙偼拏慺娷検偑崅偔側傟偽崅偔側傝傑偡偺偱丄塃偺梩孮偺梩偺傎偆偑岝崌惉懍搙偑崅偔側傝傑偡丅寢壥偲偟偰丄壓憌偺岝崌惉検偑戝偟偰曄傢傜側偄偺偵忋憌偺岝崌惉検偑堎側傞偨傔丄岡攝偑偁傞梩孮偺傎偆偑岝崌惉検偑懡偔側傝傑偡丅
丂偮傑傝拏慺暘攝棟榑偺奣梫偼埲壓偺捠傝偱偡丗庛岝偱偼丄岝崌惉懍搙偑岝偵棩懍偝傟傞偨傔丄拏慺娷検傪崅偔偟偰嵟戝岝崌惉懍搙偑崅偄梩傪嶌偭偰傕幚嵺偺岝崌惉検偼忋偑傜側偄丅偦傟偔傜偄側傜丄嫮偄岝偑摉偨傞梩偵拏慺傪傑傢偟偨傎偆偑丄偨偲偊屄梩偺岝崌惉懍搙偑掅偔側偭偨偲偟偰傕丄屄懱偺岝崌惉懍搙偼憹壛偡傞丄偲偄偆傢偗偱偡丅
丂Field (1983) 偼忋婰偺棟榑乮儔僌儔儞僕僃忔悢傪巊偭偨曽乯傪傕偲偵丄掅栘偺梩孮傪巊偭偰幚嵺偺拏慺暘攝偲丄嵟揔側拏慺暘攝傪寁嶼偟丄拏慺暘攝偺堦擔偺岝崌惉懍搙偵懳偡傞塭嬁傪挷傋傑偟偨丅寢壥偼丄岝忦審偑崅偄梩傎偳拏慺娷検偑崅偐偭偨傕偺偺丄拏慺傪嬒堦偵暘攝丒幚嵺偺暘攝丒嵟揔側暘攝傪偟偨偲偒偺屄懱偺岝崌惉懍搙傪偦傟偧傟寁嶼偟偰傒偰傕偁傑傝堘偄偑側偄丄偲偄偆丄偁傑傝僄僉僒僀僥傿儞僌偱偼側偄榖偵側偭偰偟傑偄傑偟偨丅側偍丄Field偼屄懱偺慡晹偺梩偭傁傪挷傋偨傢偗偱偼側偔丄堦偮偺姅偺拞偺俆枃乮両乯偺梩偭傁偵偮偄偰挷傋偨偩偗偱丄幚嵺偺屄懱偺岝崌惉傪寁嶼偟偨傢偗偱傕偁傝傑偣傫丅
丂Mooney傗Field偺棟榑傪梩孮岝崌惉偺棟榑偵梈崌偟偨偺偑Hirose & Werger (1987b) 偺巇帠偱偡丅斵傜偼丄傑偢丄岝-岝崌惉嬋慄傪旕捈妏憃嬋慄偱嬤帡偟傑偟偨乮旕捈妏憃嬋慄偵偮偄偰偼偙偪傜乯丅師偵奺梩偺旐捈妏憃嬋慄偺係偮偺僷儔儊乕僞朞榓壓偺嵟戝岝崌惉懍搙Pmax丄弶婜岡攝f丄嬋慄偺撌搙q丄屇媧懍搙R傪梩拏慺娷検偺娭悢偱昞偟傑偟偨乮Hirose & Werger 1987乯丅斵傜偺僨乕僞偺偐傢傝偵丄巹偑巊偭偨傕偺傪壓偺恾偵帵偟傑偡丅
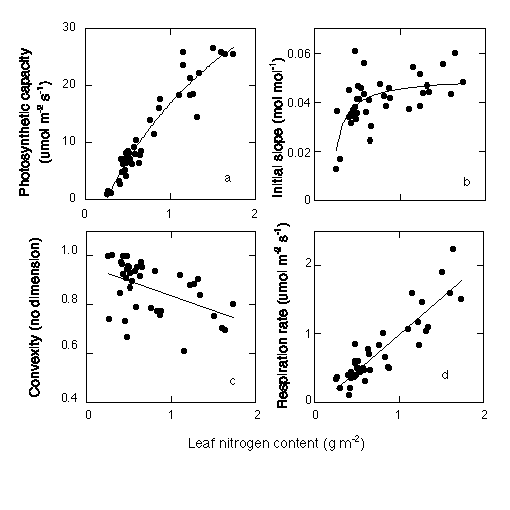
拏慺娷検leaf nitrogen content偲Pmax (photosynthetic capacity)丄f (initial slope)丄q (convexity)丄R (respiration rate)偺娭學丅僆僆僆僫儌儈偺僨乕僞丅偆偪偺尋媶幒偺廋巑壽掱傪懖嬈偟偨恵摗孨偑偲偭偨傕偺偱偡丅
偙偙偱偼夞婣幃傪偄偪偄偪帵偟傑偣傫偑丄偙傟傜偺夞婣幃傪慻傒崌傢偣傞偲堦枃偺梩偺岝亅岝崌惉嬋慄傪拏慺娷検偺娭悢偲偟偰昞偡偙偲偑偱偒傑偡丅師偵丄憌暿姞傝庢傝傪峴偄傑偡丅偙傟傑偱偺孮棊岝崌惉偺尋媶偱偼丄孮棊偺梩孮傪崅偝暿偵暘偗丄梩柺愊偲廳偝傪應傞偩偗偱偟偨偑丄斵傜偼拏慺娷検傕應掕偟傑偟偨丅偙傟偱奺憌偵偳傟偩偗拏慺偑偁傞偐傢偐傝傑偡丅Beer偺朄懃傪巊偊偽奺憌偺岝娐嫬傕傢偐傞偺偱丄拏慺娷検偲岝娐嫬偐傜丄奺憌偺岝崌惉検傪寁嶼偡傞偙偲偑偱偒傞傢偗偱偡丅埲壓偼僙僀僞僇傾儚僟僠僜僂孮棊偱摼傜傟偨愊嶼梩柺愊巜悢偲拏慺娷検偺娭學偱偡丅
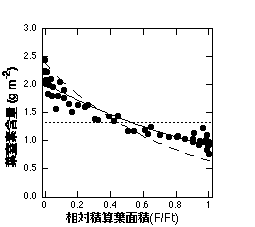
僙僀僞僇傾儚僟僠僜僂孮棊撪偺拏慺暘攝丅墶幉偼孮棊忋晹偐傜愊嶼偟偨梩柺愊傪孮棊偺憤梩柺愊偱彍偟偨傕偺偱丄0偑孮棊嵟忋晹丅仠偑幚嵺偵娤嶡偝傟偨僨乕僞丄幚慄偑N = N0 exp(-KN F/Ft)偱僼傿僢僩偟偨傕偺丄揰慄偑拏慺娷検偺暯嬒抣丄攋慄偼棟榑揑偵媮傔傜傟偨嵟揔側暘攝丅旻嶁乮1998乯傪彂偔偲偒偵峀悾偝傫偐傜惗僨乕僞傪傕傜偭偰昤偄偨傕偺偱偡丅
Hirose & Werger (1987b) 偱偼丄梩孮岝崌惉傪寁嶼偡傞偩偗偱側偔丄乽傕偟奺憌傊偺拏慺暘攝偑曄傢偭偨傜乿偲偄偆巚峫幚尡乮僔儈儏儗乕僔儑儞乯傪峴偄傑偟偨丅梩孮偑帩偮拏慺検傪堦掕偲偟丄奺憌偺拏慺娷検偑堦掕偩偭偨傜丄偁傞偄偼拏慺娷検偺岡攝傪尰幚傛傝傕媫偵偟偰傒偨傜梩孮岝崌惉懍搙偑偳偺傛偆偵曄傢傞偐傪寁嶼偟偰傒偨傢偗偱偡丅偦偺寢壥丄斵傜偑尋媶偵梡偄偨僙僀僞僇傾儚僟僠僜僂孮棊偱偼丄傕偟拏慺暘攝偵岡攝偑側偄応崌乮拏慺娷検堦掕乯偵斾傋丄幚嵺偺梩孮偺岝崌惉懍搙偼20%埲忋崅偄偙偲偑傢偐傝丄拏慺暘攝偺岡攝偵戝偒側堄枴偑偁傞偙偲偑傢偐傝傑偟偨丅傑偨丄嵟揔側梩孮偲斾傋傞偲丄拏慺岡攝偼幚嵺偺岡攝偵斾傋傗傗媫偱偟偨乮忋偺恾攋慄乯丅偟偐偟丄嵟揔側応崌偲斾傋尰幚偺梩孮偺岝崌惉懍搙偼10%埲壓偺堘偄偟偐側偔丄尰幚偺梩孮偼嵟揔偵嬤偄暘攝傪幚尰偟偰偄傞偙偲偑傢偐傝傑偟偨丅偙傟偼Field (1983) 偺寢壥偲偼懳徠揑偱偡偑丄Field偺巇帠偱偼丄柧傞偄応強偵惗偊偰偄傞掅栘傪巊偄傑偟偨偺偱丄屄懱撪偺岝娐嫬偺堘偄偑彫偝偐偭偨乮壓偺梩偱傕偁傑傝埫偔側偐偭偨乯偐傜偱偼側偄偐偲巚傢傟傑偡丅
丂偱偼嵟揔側拏慺暘攝偲偼偳偺傛偆側暘攝偱偟傚偆偐丅Farquhar (1989)偼丄嵟揔側暘攝偼丄奺憌偺庴岝検偲Pmax乮朞榓岝壓偺嵟戝岝崌惉懍搙乯偑斾椺偡傞偲偒偵梩孮岝崌惉懍搙偑嵟揔偵側傞偲巜揈偟傑偟偨丅
I'1:I'2:I'3... = Pmax1:Pmax2:Pmax3...
偙偙偱I'偼庴岝嫮搙傪丄壓晅偺悢帤偼憌偺斣崋傪堄枴偟傑偡丅偙偙偐傜嵟揔側拏慺暘攝傪寁嶼偱偒傑偡丅Anten et al. (1995a) 偼丄Pmax偲拏慺娷検乮Narea乯偑Pmax=a(Narea-b)偲乮a偼孹偒丄b偼x愗曅乯捈慄偺娭學偵偁傞偲偒丄嵟揔側拏慺暘攝偼埲壓偺傛偆偵側傞偙偲傪帵偟傑偟偨丅
丂
Narea - b = (NT-bFT) K exp(-KF)/(1-exp(-KFT))
丂
偙偙偱NT偼梩孮慡懱偺拏慺娷検乮搚抧柺愊偁偨傝乯丄K偼媧岝學悢丄F偼愊嶼梩柺愊巜悢丄FT偼憤梩柺愊巜悢偱偡丅偨偩偟丄偙偺幃偑惉傝棫偮偵偼偄偔偮偐忦審偑偁傝丄弶婜岡攝f傗嬋慄偺撌搙q偑堦掕偱偁傞偙偲丄拏慺娷検偲屇媧懍搙R偑斾椺偟偰偄傞側偳偑昁梫偱偡丅傗傗偙偟偄幃偱偡偑丄偙偪傜偵彂偄偨傛偆偵丄K exp(-KF)偑憌偺庴岝検傪堄枴偟傑偡丅(NT-bFT)/(1-exp(-KFT))偼掕悢側偺偱丄Narea - b偼庴岝検偲斾椺偡傞偙偲偵側傝傑偡丅Pmax=a(Narea-b)偱偡偐傜丄梫偼丄忋偵傕彂偄偨傛偆偵庴岝検偲Pmax偑斾椺偡傞傛偆偵拏慺傪暘攝偡傞偙偲偑嵟揔偩偲偄偆偙偲偱偡丅偳偆偟偰偙偆側傞偐丄悢妛揑側徹柧偼尨挊傪偛棗偔偩偝偄丅
丂偝偰丄偙偺幃偵傛傝丄嵟揔側拏慺暘攝傪斾妑揑娙扨偵寁嶼偱偒傞傛偆偵側傝傑偟偨乮偦傟傑偱偼僔儈儏儗乕僔儑儞偱悢抣寁嶼偟側偄偲嵟揔暘攝偑弌側偐偭偨偺偱偡丅Hikosaka and Hirose 1997偱偼嬯楯偟傑偟偨乯丅尰幚偺拏慺暘攝偼嵟揔暘攝偐傜偳傟偩偗棧傟偰偄傞偺偱偟傚偆偐丅Anten et al. (1995) 偱偼埲壓偺幃偱拏慺暘攝傪夞婣偟丄夞婣學悢KN傪摼傑偟偨丅
丂
Narea - b = (NT-bFT) KN exp(-KNF)/(1-exp(-KNFT))
丂
偙偺幃偼K偑KN偵曄傢偭偨偩偗偱偡丅偮傑傝KN偲K偑摍偟偗傟偽嵟揔偲偄偆偙偲偵側傝傑偡丅偱丄條乆側梩孮偺KN偲K傪斾傋偰傒傞偲丄KN偲K偺娫偵偼崅偄憡娭偑偁傝傑偟偨偑丄悢帤傪尒傞偲丄KN偼K偺敿暘掱搙偱偟偨乮Anten et al. 2000乯丅偮傑傝尰幚偺暘攝偼嵟揔側暘攝傛傝傕偐側傝娚傗偐偱偁傞偲偄偆偙偲偱偡丅拏慺暘攝偑忢偵嵟揔傛傝娚傗偐偱偁傞偙偲偼堄枴偑偁傝偦偆偱偡丅岡攝傪媫偵偡傞偲偄偆偙偲偼嵟忋晹偺拏慺娷検傪堎忢偵崅傔傞偙偲偵側傝傑偡偑丄偙傟偼怉怘幰偵桳棙偱偁傞偺偱怉暔偵偲偭偰偼晄棙偱偁傞乮Stockhoff 1994乯丄偲偄偆峫偊曽偑偁傝傑偡偑丄幚嵺偺偲偙傠偼傑偩傛偔傢偐偭偰偄傑偣傫丅
丂
丂忋偱偼拏慺傪峫椂偟偰偄側偄忬懺偱偺嵟揔梩柺愊巜悢偺榖傪偟傑偟偨丅岝崌惉偵懳偡傞拏慺偺栶妱傪峫偊傞偲丄偳偺梩柺愊偑嵟揔偐偑曄傢偭偰偒傑偡丅
丂拏慺偲乽嵟揔梩柺愊巜悢乿偺娭學偵偍偄偰偼丄拏慺偑桳尷偺帒尮偱偁傞偙偲偲偄偆偙偲偑嵟傕廳梫側栤戣偱偡丅搚忞偺拏慺塰梴偑怉暔偺惗堢傪棩懍偡傞偙偲偼丄擾嬈偵偍偄偰傕帺慠奅偵偍偄偰傕傛偔偁傞偙偲偱偡丅孮棊乮屄懱乯偑帩偮拏慺偑尷傜傟偰偄傞偲峫偊傑偟傚偆丅拏慺偑彮側偄偺偵偨偔偝傫偺梩柺愊傪帩偮偲丄屄乆偺梩偺柺愊偁偨傝偺拏慺擹搙偑壓偑傝傑偡丅偦偆偡傞偲丄屄乆偺梩偺Pmax偑掅偔側傝丄嫮偄岝偑摉偨偭偰傕偨偄偟偰岝崌惉偱偒側偄丄偲偄偆忬懺偵側傝傑偡丅媡偵梩柺愊偁偨傝偺拏慺擹搙傪忋偘傞偨傔偵梩柺愊傪尭傜偟偡偓傞偲丄梩偑彮側偄偨傔偵岝傪廩暘媧廂偱偒側偔側傝傑偡乮壓恾乯丅偲偄偆偙偲偱丄梩孮偑拏慺傪偳傟偩偗帩偭偰偄傞偐偱嵟揔側梩柺愊巜悢偑堎側傝丄摨偠媧岝學悢偱傕帩偭偰偄傞拏慺偑彮側偄傎偳嵟揔梩柺愊巜悢偑壓偑傝傑偡乮Anten et al. 1995b乯丅
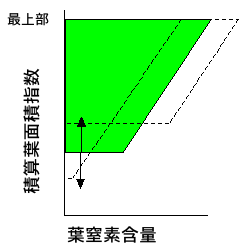
丂偙偺懠丄偳偪傜偐偲偄偆偲嵶偐偄栤戣側偺偱偁傑傝巜揈偝傟傞偙偲偼偁傝傑偣傫偑丄拏慺傪峫椂偡傞偲媧岝學悢偲嵟揔梩柺愊巜悢偺娭學偑偑傜偭偲偐傢傝傑偡丅Monsi-Saeki儌僨儖偱偼梩偺Pmax傪堦掕偵偟偰偄傑偟偨丅偦偺偨傔丄媧岝學悢偑戝偒偄梩偱偼丄庴岝検偑崅偄傢傝偵岝崌惉懍搙偑掅偔丄岝傪媧廂偟偰傕岝崌惉偵巊偊側偄丄偲偄偆忬懺偵側偭偰偟傑偄傑偟偨丅偙偺偨傔丄梩柺愊傪嵟揔壔偟偨偲偒偺岝崌惉懍搙偑媧岝學悢偑戝偒偄傎偳掅偔側傞傢偗偱偡丅偟偐偟丄拏慺暘攝傪峫椂偟丄忋晹偺梩偵偨偔偝傫拏慺傪暘攝偱偒傞偲側傞偲榖偼堘偄傑偡丅嫮偄岝傪庴偗傞梩偼崅偄Pmax傪帩偮傢偗偱偡丅壖偵丄庴岝検偲Pmax傪姰慡偵斾椺娭學偵偍偗傞偲偡傞偲丄Monsi-Saeki儌僨儖偱偁偭偨傛偆側柍懯偼慡偔側偔側傝丄孮棊岝崌惉懍搙偼孮棊偺岝媧廂検偵斾椺偡傞偙偲偵側傝傑偡丅摨偠梩柺愊巜悢乮嵟揔梩柺愊巜悢傛傝掅偄応崌乯偱斾妑偡傞偲丄孮棊岝崌惉懍搙偼媧岝學悢偑崅偄傎偳崅偔側傝傑偡丅偙傟偼Monsi-Saeki儌僨儖偱傕摨偠寢壥偱偡偑丄嵟揔梩柺愊巜悢偵偍偗傞孮棊岝崌惉懍搙偼堎側傝丄媧岝學悢偑崅偄傎偳崅偄偲偄偆寢壥偑偱傑偡乮Anten et al. 1995b乯丅
丂偟偐偟丄偙傟偼屄梩偺拏慺娷検偑偳偙傑偱傕崅偔側傟傞偙偲傪壖掕偟偨応崌偱偡丅偦偺傛偆側乽棟憐揑側乿拏慺暘攝偵偡傞偲丄嵟忋晹偺梩偺拏慺擹搙偼300 mmol N m-2 偲偐偄偆偁傑傝尰幚揑偱偼側偄抣傪帩偮偙偲偵側傝傑偡乮Anten et al. 1995a. 斵偼丄慡偔偁傝摼側偄抣偱偼側偄丄偲庡挘偟偰傑偡偑乯丅幚嵺偵偼丄尰幚偺孮棊偺拏慺暘攝偼悢妛揑偵摫偐傟傞嵟揔側暘攝偲偼堘偄傑偡丅傑偨丄Anten et al.偺儌僨儖偱偼丄拏慺娷検偲Pmax偺娭學傪捈慄偲偟偰偄傑偡偑丄惗棟妛揑偵尒偰傕丄拏慺傪憹傗偣偽Pmax偼嵺尷側偔忋偑傞偐丄偲偄偆偲丄偦偆偲傕巚偊傑偣傫丅CO2奼嶶偺栤戣側偳傕偁傝丄偁傞掱搙拏慺娷検偑憹偊傞偲Pmax偺憹偊偼摢懪偪偵側傞偲巚傢傟傑偡丅尰幚偵丄偄偔偮偐偺庬偱偼拏慺娷検偲Pmax偺娭學偑摢懪偪偵側傝傑偡乮忋偺恾嶲徠乯丅偙偺俀揰偐傜丄傗偼傝梩孮偺嵟戝岝崌惉懍搙偼媧岝學悢偑掅偄傎偆偑崅偄丄偲偄偆偺偑尰幚揑偩偲巚偄傑偡丅
丂拏慺娷検偲Pmax偺娫偺憡娭偑朞榓宆偺嬋慄乮摢懪偪乯偱偁傞偲壖掕偡傞偲丄嵟揔側暘攝傗梩柺愊巜悢偑丄尰幚偺梩孮偲偦偆曄傢傜側偄抣偵側傝傑偡丅壓偺恾偼Hikosaka & Hirose (1997) 偵嵹偣懝側偭偨傕偺乮惓偟偔側偄恾偑嵹偭偰偄傑偡乯偱偡丅
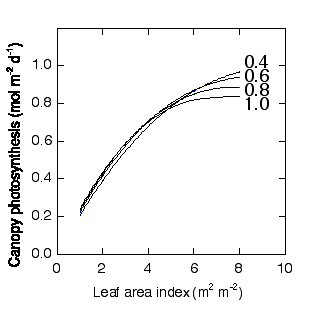
廲幉偑梩孮岝崌惉懍搙丄墶幉偑梩柺愊巜悢丄恾拞偺悢帤偼梩孮偺媧岝學悢乮K乯傪帵偟傑偡丅
偙偙偱偼丄孮棊偺拏慺検傪堦掕偵偡傞偺偱偼側偔丄孮棊偺暯嬒梩拏慺娷検偑堦掕偵側傞偲偄偆壖掕傪偍偄偰偄傑偡丅拏慺暘攝偼嵟揔壔偝傟偰偄傑偡丅偙偙偱巊偭偰偄傞拏慺偲岝崌惉偺娭學偼Hirose & Werger (1987a) 偵傛傞僙僀僞僇傾儚僟僠僜僂偺僨乕僞偱偡丅偙偙偱偼拏慺娷検偲Pmax偺娫偼捈慄偱偁傞偲壖掕偝傟偰偄傑偡偑丄嬋慄偺撌搙乮q乯偑崅拏慺娷検傎偳壓偑傞偲偄偆壖掕偑偍偐傟偰偄傑偡丅偙偺壖掕偼丄幚幙忋拏慺偲Pmax偺娭學偑嬋慄偱偁傞偙偲偲摨偠堄枴傪帩偪傑偡丅偙偆偄偭偨壖掕傪偍偔偲丄孮棊岝崌惉懍搙偺嵟戝抣偼媧岝學悢偑崅偄傎偳掅偔側傝傑偡丅
丂傑偨丄Hirose et al. (1997) 偱偼丄孮棊拏慺検丒梩柺愊巜悢丒孮棊岝崌惉懍搙偺娭學傪僔儈儏儗乕僔儑儞偱挷傋傑偟偨丅偙偺僔儈儏儗乕僔儑儞偵傛偭偰傢偐偭偨偙偲偼丄嵟揔側孮棊拏慺検偑偁傞偲偄偆偙偲偱偡丅梩屇媧懍搙偲梩拏慺娷検偑斾椺偡傞偲偄偆壖掕傪偍偄偰偄傞偺偱丄孮棊偺拏慺検偑懡偡偓傞偲偐偊偭偰孮棊岝崌惉懍搙偑掅偔側傞傢偗偱偡丅
丂側偍丄拏慺傪峫椂偵擖傟偨応崌偼丄嵟壓憌偺梩偺岝曗彏揰偼傎偲傫偳栤戣偵偼側傜側偔側傝傑偡丅尰嵼巊傢傟偰偄傞孮棊岝崌惉儌僨儖偱偼丄屄梩偺屇媧懍搙偼拏慺娷検偲惓偺憡娭傪帩偪傑偡丅偙偆偄偆壖掕傪偍偔偲丄嵟壓憌偺拏慺娷検傪壓偘偰屇媧懍搙傪壓偘傞偲丄偦傟偲摨偠暘偩偗忋憌偺梩偺屇媧懍搙偑忋偑傝傑偡丅偟偨偑偭偰丄孮棊偺屇媧懍搙偼孮棊偺帩偮拏慺検偵埶懚偡傞偙偲偵側傝丄屄梩偺屇媧懍搙傪峫偊傞昁梫偑側偔側傞傢偗偱偡乮Anten et al. 1995乯丅