光合成の生理生態学講座
目次
はじめに (200706 13)
太陽放射 (200706 13)
直達光と散乱光 (200707 15)
サンフレック (200007 06)
群落内における直達光と散乱光の分布 (200707 30)
個葉の受光(200708 07)
光の質(200708 07)
光合成のエネルギー源は光です。野外の光環境は時間・空間的に大きく異なり、光合成速度の時間・空間的変動を引き起こします。ここでは植物の光合成から見た光環境について書きます。特に、植物群落内部の光環境について詳しく書きたいと思います。
我々が通常「光」と呼ぶのは、太陽が放射する電磁波のうち400~700nmの波長域の電磁波のことをいいます。これは我々の目で見ることができることから可視光と呼ばれます。光合成で利用できる光も400~700nmに限られます。400~700nmの光のエネルギーを光合成有効放射(Photosynthetically active radiation、PAR)と呼びます。これは「放射」なので通常W m-2という単位で表されます。
地球が大気圏外で太陽から受ける放射は、全波長を合わせると約1370 W m-2です(太陽光に対して垂直な面が受ける場合)。このうち一部は大気中の物質に吸収されたり散乱させられたりし、地表面に到達するのは約1000 W m-2です。このうちPARは約半分の500 W m-2程度です。
一粒の光量子がもつエネルギーは波長によって異なり、短波長の光ほど光量子あたりのエネルギーは大きくなります。しかし光合成は光量子のエネルギーには依存せず、光量子が何個吸収されたかが重要です。このため光合成研究では光の強さがエネルギーで表されることはほとんどなく、光量子束密度(photon flux density、PFD)あるいは光量子フラックス(photon flux、PF)として表されます。400~700nmの光量子束密度を光合成光量子束密度(Photosynthetic photon flux density、PPFD)と呼びます。余談ですが、最近はPFDではなくPFが用いられるようになってきています。flux自体に「面積あたりの」という意味があるため「密度」は不要である、という理由なのですが、私は個体あたりの光吸収量とか「面積あたり」を含まない意味の光量子量を使ったりしているので、PFよりはPFDのほうが好きです。
地表面が受ける太陽放射は時間と場所によって大きく変動します。これは主として地球が球体であることと地球の自転・公転によって時間・場所によって太陽高度が異なること最も重要です。太陽高度が高いほど地表面は太陽光に対して垂直に位置することになり、面積あたり受ける放射が大きいことになります。一日の光強度(I)の変化はサイン二乗カーブで近似することができます(Hirose and Werger 1987a)。
I = Ioo sin2{π(t-6)/12} (6 < t < 18)
I = 0 (0 < t < 6, 18 < t < 24)
ここで、tは一日の時間(hour)、Iooは太陽が南中した時点(t=12)の光強度です。日本の緯度では、快晴時にIooはPPFDで2000μmol m-2 s-1を超えるのが普通です。この式では、6時から18時まではサイン二乗カーブに則って光環境が変化し、それ以外は真っ暗、という仮定をしています(春分・秋分の日の仮定です)。式をうまくいじると昼夜の長さを変えることができます(Hirose et al. 1997)。
もう一つ重要な要素は大気の状態です。雲が出ると太陽光が遮蔽され、地表に届く放射が減ります。大気中のちりなどが増えても散乱・吸収が起こるため地表に届く放射が減ります。
野外において植物が受ける光は、直達光と散乱光の二種類に分けることができます。直達光は太陽から直接届く光で、散乱光はそれ以外の光です。散乱光ももとはといえば太陽からくる光に由来します。直達光が大気中のちりなどにより方向を変えられ(=散乱)、太陽がない方向から光が届くわけです。ちなみに大気がない宇宙空間では散乱はほとんど起きず、例えば月では地表面に届く光のほとんどは直達光で、太陽がある部分以外の空はほとんど真っ黒、つまり散乱光はほとんどありません。
地表面に届く散乱光と直達光の割合は条件によって大きく異なります。例えば、曇天の日は届く光の全てが散乱光となります。また、黄砂など大気中のちりが多いときも散乱光の割合が大きくなります。晴れた日でも朝・夕は散乱光の割合が大きくなります。直達光の割合が最も高い条件は快晴で太陽高度が最大になったときです。快晴ならば、植物が一日に受ける光のうち1/4が散乱光なのだそうです(Gates 1980)。日本語ではCampbell and Norman (1998) の和訳(生物環境物理学の基礎)をご覧ください。
植物群落内の光環境は非常に複雑です。多くの植物群落内部では、地面に当たる光の強さは群落最上部にあたる光より二桁小さくなります。このような光環境の不均一性は植物群落の生産力に大きな影響を及ぼします。
さて、森林でも草原でも群落の内部は暗い、というのは常識だと思います。上部にある葉によって光が吸収されてしまうからです。次に、光が遮られる、ということを考えてみましょう。まず単純な例から考えます。ある液体があるとしましょう。この液体中をある波長の光が1cm進むと、強度が半分になってしまうと考えましょう(つまり、半分は吸収されてしまう)。当てる光を1としたとき、液体中をx cm進むと光強度はどれだけになるでしょうか? 答は、0.5xです。半分がまた半分になり、それがまた半分になり・・・を繰り返すわけです。光がある確率で減衰していく場合、光強度は距離に対し直線的ではなく、指数関数的に減衰していきます。これがBeerの法則です。具体的には、距離Xと光強度Iの関係は、
I = Io exp(-KX) (1)
という関係にあります。ここで、Ioは最初の光強度、Kが吸光定数とか減衰定数とか呼ばれるものです。Kが大きいほど、液体の吸収能力が高いことになります。群落内の場合は、葉っぱが光を吸収します。葉っぱが多いほど光の減衰が激しくなります。上と同様な式で表されます。
I = Io exp(-KF) (2)
ここでFは積算葉面積と呼ばれるものです。土地面積あたり存在する葉面積を上部から積算していったものです。具体的には、群落を高さごとに層別にわけ、各層の土地面積あたりの葉面積を上から順に積算していったものです。ある層で測定した光強度を考える場合、その測定した位置より上にある葉の面積と光強度の間に上の式の関係が成り立ちます。ちなみに、その群落の全ての葉面積を土地面積で割った値を葉面積指数(leaf area index)と呼びます。この分野では(しばしば何の説明もなく)LAIと略されます。
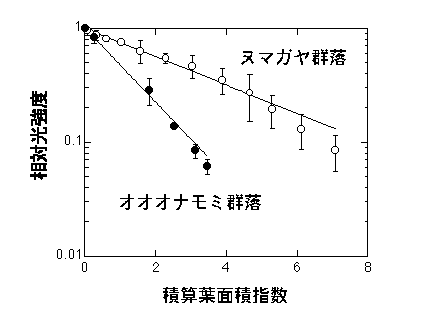
ヌマガヤ群落(葉が垂直的)とオオオナモミ群落(葉が水平的)における相対光強度(I/Io)と積算葉面積指数Fの関係。ヌマガヤ群落のKは0.29、オオオナモミ群落のKは0.75。
この関係はMonsi & Saeki (1953)によって実験・理論の両面から明らかにされました。数学的には、Beerの法則と無関係に上の式を理論的に導くことができます。詳しくは、「植物の物質生産」(ボイセンイェンセン著・野本訳)の最後の部分に載せられている、Monsi & Saeki (1953)の日本語訳をご覧下さい。注意していただきたいのは、群落の下に届く光、というのは、「葉を透過してくる光」と「葉のない部分を通ってくる光」の和である、ということです。葉の光透過率は5-10%程度です。積算葉面積がそれほど高くない層(土地面積あたりで3以下)だと、「葉のない部分を通ってくる光」がほとんどです。ときどき、葉を透過してくる光がほとんどだと考える人がいますが、そうではありません。理論的な仕事では、葉が空間内にランダムに散らばっていると考え、その中で光が遮られる確率を考えると上の式を導くことができます。
さて、吸光係数Kについてです。液体の話同様、Kが大きいことは光が遮られやすいことを意味します。理論的には、Kには以下のように様々な要因が影響を与えます。
1 葉の角度
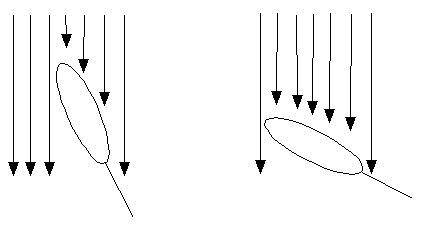
簡単に想像できると思いますが、葉が垂直的に立っているほど光は下に届きやすくなります(上の図も参照)。
2 葉の大きさ
理論的には、一枚一枚の個葉の面積が大きいほど吸光係数は大きくなるはずです。小さい葉っぱがたくさんある場合のほうが隙間を光が通過する確率が高く、Kが小さくなります。
3 葉の空間分布
葉の空間分布はKに大きな影響を与えます。まず、水平的に集中分布している場合を考えてみましょう。例えばセイタカアワダチソウのように、一本のひょろ長いまっすぐな茎に、葉柄のない小さな葉がたくさんついているところを想像してみて下さい。一本一本の植物は細い円筒のようになっていて、各個体の間には距離があるとします。ある水平面で葉の断面を見てみると、葉が集中しているところと、葉がほとんどない部分とに分かれることになります。このとき、葉がランダムに分布している群落に比べ、光は下に届きやすくなり、Kは小さくなります。次に、垂直的に集中分布している場合を考えてみましょう。水平断面ではランダムだけど、垂直断面では集中分布が見られる場合です。このときは、光はもれにくくなります。もし垂直断面で一点に葉が集まると、ほとんど光は下に漏れない、ということを想像できると納得いくのではないかと思われますが・・・。
しかし、葉っぱの大きさがKに有意な影響を与えるのは例えばフキみたいに極端にでかい葉っぱの場合のみで、空間分布でも、かなり強い集中分布が起こらない限りはそれほどKは変わりません。というより、一般に見られるような植物では葉の空間分布はそれほどランダムから離れていないし、大きさも充分小さいのです。したがって、現実的には、Kの大きさに最も影響があるのは、葉の角度である、ということになります。Monsi & Saeki (1953) は葉の角度とKの関係を理論と実測の両面から示しています。葉の角度と吸光係数の理論的関係は下で説明します。
上記Beerで表されるIは、ある高さにおける光強度の期待値であって、実際にどこでもその光強度になるわけではありません。すでに書いたように植物が受ける光には直達光と散乱光があります。ある高さにおける散乱光の強度は比較的一定ですが(それでもばらつきがないわけではない)、直達光の場合は、測定点と太陽の間にたまたま遮蔽物(上層の葉とか)があるか否かで大きく光強度が変わります。例えば森林の林床では散乱光が届く光強度は最上層の数%にすぎませんが、直達光の場合はもし届けば1000μmol m-2 s-1の光が突如あたることになります。このような強い光をサンフレック(sunfleck)とよびます。木漏れ日とか陽班とか近い日本語はあるのですが、どれも微妙に違うので、このページでは以下サンフレックと綴ることにします。
サンフレックが届くか否かは太陽の動きと上層の葉の位置に依存するため、空間的にも時間的にも変動することになります。ハワイの森林の林床がどのような光環境かを調べた仕事を少し紹介しましょう(Pearcy and Calkin 1983, Pearcy 1983)。Pearcyらが調べた林床では、直達光があたらないときは葉が受ける光強度はわずか数10 μmol m-2 s-1です。しかし、ときおり強い光があたります。この葉が一日に受ける光は 86 μmol cm-2 d-1ですが、このうち40%はサンフレックによるものです。このサンフレックは、ほとんどが250 μmol m-2 s-1以上の光強度で、ものによっては1500 μmol m-2 s-1を超えます。ただ、長さは短く、サンフレックの2/3は30秒以内しか続きません。
もちろんこれは一例にすぎません。様々な群落でどのようにサンフレックが当たるかが調べられていると思いますが、私はよく知りません。とりあえず、ダイズ群落(Pearcy et al. 1990)、ススキ群落(Tang et al. 1992)を挙げておきます。
森林の林床でも、ギャップがあると時間の長い直達光を浴びることになります。これは「長いサンフレック」ということになりますが、こういう長いのは「sun-patch」と呼んでサンフレックと区別しよう、と提唱している人がいるらしいですが、私は知りません(寺島さんに昔聞いた話です)。
上記のようにBeer則はある高さの光強度の期待値を出すもので、実際にはある高さにおける光強度は直達光があたっているか否かで大きくことなります。また、散乱光はあらゆる方向から降り注ぎます。さらに、特に草本群落では、葉群内に角度の異なる葉が混在していることは珍しいことではありません。このような複雑な状況は、上記のBeer則で表した場合には、ある意味平均化されてしまうことになります。しかし現在では、「葉が充分小さくランダムに分布している」という仮定があればこのような複雑な状況も数式で表現できます。ここではそれぞれの要因がどのように扱われるのか、単純化した直観的な説明をします。詳しい数式の説明は、公表されている最新のものとしてAnten (1997) をごらんください。現在準備中の我々の論文が受理されましたら詳しいものをここに掲載しようと思います(いつになるやら?)。
・直達光の減衰
直達光の場合、光強度は減衰しないと仮定します。そのかわり、上部の葉の遮蔽によって内部へ入っていく光の束の数が減っていくと考えます。具体的には、ある高さにおいて直達光があたる葉の割合を計算によって求めます。この割合が群落下部ほど少ない、という計算をするわけです。
この計算では葉は「直達光が当たる部分」と「直達光が当たらない部分」に二分されますが、これはあくまでも近似です。太陽は天球角度で約1度の大きさがあるため、太陽の一部が上部の遮蔽物に隠されてしまうような状況が存在します。したがって「直達光があたる部分」と「あたらない部分」の間には文字通りグレーゾーンが存在します。しかしこのグレーゾーンの大きさは遮蔽物と葉の間の距離に依存するなど面倒なことが多いため、モデルに組み込むことは用意ではありません(考慮に入れた光減衰モデルの存在は聞いたことがありません)。
また、太陽の移動を考慮する必要があります。太陽は東から昇り、南中して西に沈むわけですが、太陽の移動とともに葉群への入射光の角度が変わることになります。葉群内の葉の角度が一定である場合、上部から入射した光と斜め方向から入射した光の減衰のしかたは異なります。例えば葉が水平であった場合は上部からの光は減衰しやすいですが、水平に近い方向からの光は減衰しにくく、葉群の奥まで届くようになります。葉が垂直的である場合はまた異なります。計算においてはこのことを考慮します(下参照)。
・散乱光の減衰
散乱光は常時様々な方向から降り注ぎます。そして、入射角によって光の減衰の仕方が異なることは直達光と同様です。このため散乱光を入射角によって区分し、各角度からの光がどう減衰するかをそれぞれ計算し、ある層における入射光量をその和として計算することになります。
直達光の減衰は、上記のように、群落の下部でも光強度は変化せず、そのかわり直達光があたる葉の割合の減少として記述されますが、散乱光の場合は全ての光が受光し、光強度が減衰するように記述されます。散乱光はあらゆる方向から降り注ぐため、ある方向の散乱光を受けない葉でも別の方向の散乱光を受ける可能性があるからです。
以上の計算の結果、ある層には直達光と散乱光を受ける葉と散乱光のみ受ける葉が混在することになります。
・様々な角度の葉が混在している場合
もし様々な角度の葉の分布がランダムであると仮定できるならば、光の減衰は平均値的な吸光係数の値を使って近似することができます。
Idir=Iodir exp (-Kdir F)
下付のdirは直達光を意味します。
直達光の吸光係数は以下の式で計算することができます。
Kdir = Oav/sin(Bs)
ここでOavは太陽の入射角に対する葉の平均投影面積割合を、Bsは太陽高度(水平面からの太陽高度入射角度)を表します(Oavについては下でも説明します)。群落内の葉があらゆる方位を向いているとすると、Oavは以下の式から求められます。
Oav = sin(Bs) cos(BL) Bs>BL
Oav = (sin(Bs) cos (BL) arcsin(tanBs/tanBL) + (sin^2(BL)-sin^2(Bs))^0.5 Bs<BL
ここでBLは葉の水平面からの傾斜角度を表します。上の式は葉の表側のみに光が当たる場合、下の式は葉の裏側にも光があたる(太陽が傾いている)場合を意味します。Oavは葉の傾きによって値が異なります。葉群全体の値を出すためには、角度の異なる葉ごとにOavを計算し、角度の頻度を考慮した上、Oavの中央値をもって葉群のOavの近似値として使用します。
散乱光も原理的には同様の計算によって得られます。散乱光の場合は、天球を高度(水平面からの角度)ごとに分け、各角度の入射光ごとに減衰曲線を計算します。入射角と葉の角度は、直達光の計算同様に考慮されます。
簡単にかつわかりやすく説明するのはなかなか難しいです。詳しくはAnten (1997) をごらんください。
葉がどれだけ受光しているかを調べるために、光センサを葉の上や横に置き、葉が受ける光強度を推定することがあります。ここで何も考えずに光強度を測定すると失敗します。葉の角度の影響が大きいためです。
光が天頂から降り注ぐ場合、葉や光センサが受ける光強度は水平面で受光したときに最大になります。葉が傾いていると、葉が受けることができる光の量が減るため、葉面積あたりの受光量が減ります。
光センサの値も同様に減ります。どの程度受光量が減るかは葉やセンサの傾きに依存します。葉の水平面からの傾斜角度をBLとすると、降り注いできた光強度Iと受光量I'の関係は
I'=I cos(BL)
となります。これはコサイン則と呼ばれているそうです(名前をつけるほどのものとも思わないが・・・)。この式は実は上のOavの式と同じものです(光が天頂からくるということはつまりsin(Bs)=1)。上記のOavは、ようするにI'/I、葉が光源に対し垂直だった場合に比べたときの受光強度を表す値です。
葉群内でも、ある層における光強度Iと実際にその層の葉が受光した光の量は異なります。ある高さnで測定した光強度をIn、その下にある高さn+1で測定した光強度をIn+1とすると、nからn+1の間で吸収された光強度はIn-In+1ということになります。葉面積あたりの受光量Iabsは、In-In+1を、この層に存在する葉面積(Fn+1-Fn)で割ればいいことになります。
Iabsn= (In-In+1)/(Fn+1-Fn)
これは、(Fn+1-Fn)→0とするとBeer則を微分することに相当します。
Iabsn= -d(Io exp(-KF))/dF = K Io exp(-KF) = K In
ということで光強度に吸光係数Kをかけたものが葉面積あたりの受光量に相当することになります。これは上記Oavの式とそれに続く式で言っていることとだいたい同じです。
ここでいうIabsは葉が吸収した光です。これは葉にあたった光と同じではありません。例えば、いわゆる光-光合成曲線の光強度は、普通葉にあたった光の強度のことですから、Iabsをそのまま光合成速度の推定に使うことはできません。葉はあたった光のうち5~10%を透過します。透過率をmとすると、
Iinc=Iabs/(1-m)
などの式で葉に当たった光強度Iincを推定することができます。
ここで言う光の質とは、ようするに光の波長組成のことです。クロロフィルは青と赤に吸収極大があり、この波長の光をよく吸収します。一方、緑の光は赤や青に比べるとあまり吸収されませんし、近赤光(700nm以上の光。赤外線のうち波長が短いもの)はほとんど吸収されません。このため、葉群下部の光波長組成は、近赤光は葉群上部とそれほど変わらないのに対し、可視光は非常に光強度が弱く、また、可視光のうちでも赤や青に比べ緑の光がやや多くなります。
植物はこのような波長組成の変化を利用しています。ファイトクロム(フィトクロムphytochrome)とよばれる色素は近赤光を吸収するか赤色光を吸収するかで分子の立体構造が変化します。近赤光と赤色光の比によって両者の存在比が異なり、これをシグナルとして自分の周囲の光環境変化を感知しています。近赤光の比率が高まることはすなわち周囲に植物が存在していることを意味し、被陰されないよう茎の伸長速度を増加させるなどの応答が知られています。