光合成の生理生態学講座
はじめに(200612 27)
コストとベネフィット(200612 27)
光合成能力のコストとベネフィット(199806・200612 27)
最適な気孔コンダクタンス(200001 10・200612 27)
定義(200205 17)
窒素生産力と平均窒素滞留時間の積としての窒素利用効率(200206 11)
窒素生産力と平均窒素滞在時間(200206 21)
窒素利用効率を考える意義(200207 07)
定義(200208 11)
水利用効率に影響を与える要因(200208 11)
安定同位体の利用(200208 22)
水利用効率の意義(200209 02)
コスト-ベネフィットと資源利用効率の考え方(200207 20・200208 01・200612 27)
植物のふるまいを進化生態学的にとらえる、すなわち、どのような戦略が進化的に有利なのか、という観点で研究するときに、私たちは、より速く成長したものが有利であると考えがちです。しかし、野外の群落を見ると、例えばいわゆる極相種と呼ばれる種は多くの場合成長速度が非常に遅い種です。速く成長する植物が優占していることはまれで、このような単純な考え方では植物の戦略を理解できないことがわかります。植物が成長するためには、例えば光、水、栄養塩といった様々な資源が必要です。ここでいう資源とは、成長に必須で、かつ供給が有限で、消費されてしまうものをいいます。一般に、自然環境では、全ての資源が充分に存在するという状況は極めてまれです。ある資源が不足しがちな場合、成長を速くするよりも、資源を効率よく利用する戦略のほうが重要かもしれません。このような観点から、資源利用効率を初めとする様々な考え方が提案されています。ここではこれらの考え方を紹介します。
まず我々が考えなければいけないのは、ある性質(戦略)についての「有利」「不利」の定義です。野外の植物を扱う場合には、進化生態学的には「適応度を高めるような性質」が有利である、と定義することができます。ここで適応度とは、その個体が残す子孫の和です。したがって、「ある性質が適応的である」というためには、その性質をもつ個体ともたない個体に子供を産ませ、その数を数えなくてはいけません。しかしそのような仕事は必ずしも容易ではありません。特に光合成などのように器官レベルの現象の場合、個別の性質と子孫生産の間に様々な要因が介在するため、目的とする性質の直接的な影響を評価することはなかなか難しいことです。
そこで、目的とする性質による「利益」と「実現するために必要なコスト」を定量化することにより、経済学的な観点から評価するということが行われています。何かの利益を得るためには、何らかのコストを払う必要があります。光合成で考えると、例えば、光合成をするためには、光合成系のタンパク質を作る、というコストがかかります。このときには、タンパク質の材料となる窒素や、タンパク質を合成するためのエネルギーが必要となります。あるいは、葉っぱを作るための材料やエネルギーが必要です。このときの、コストと利益の関係を「適応性」の判断基準に使います。ある利益を得るためにかかるコストを最小にできたものが「適応的に有利である」と考えられます。少なくとも、コストを最小にできなかったものよりは有利だよ、ということです。
しかし、「コスト」も様々です。条件によって炭素だったり、光だったり、窒素だったり、水だったりします。なぜ「コストが多様か」についてわかりやすい例を挙げましょう。題して「自動車のコストとベネフィット」です。経済速度というのがあります。1リットルのガソリンで最も長く走れる距離です。燃費(1リットルあたりの走行距離)というのは速度に依存します。例を挙げるまでもないかもしれませんが、亀のような速度で走っているときと40km/hの速度では、後者のほうが燃費がいいです。同様に、もっと高速(例えば200km/h)で走ると、かえって燃費が下がります。ここでは、経済速度を40kmとしましょう。さて、学生は一般的に貧乏ですが、有り余る時間を持っています。逆に社会人の金持ちは忙しいと考えましょう。学生と、社会人では、車の「適応的な」運転のしかたが違います。貧乏人はガソリンが買えません。したがって、ガソリンを効率よく使うことが重要です。幸い彼には時間がありますから、40kmでちんたら走るのが彼にとって最も有利な走り方です。一方、社会人は時間がありません。幸い金はありますから、ガソリンを大量に使ってでも高速で車を運転することが有利になります。植物の場合は、窒素が足りなければ窒素を、水が足りなければ水を効率よく使うことが「適応的」である、ということになります。
さて、コストとベネフィットを定量化したとして、どのような戦略が有利かを判断するにも複数の方法があります。一つは、ベネフィットとコストの差を最大にする戦略が最適であるという考え方です。もう一つが、ベネフィットとコストの比を最大にする戦略が最適であるという考え方です。獲得(あるいは保有)資源あたりのベネフィットは資源利用効率と定義され、多くの研究で用いられています。以下からは、光合成に関するコスト-ベネフィットの使用例をいくつか紹介します。「光合成能力のコストとベネフィット」、「窒素利用効率」、「水利用効率」はベネフィットとコストの比を、「最適な気孔コンダクタンス」ではベネフィットとコストの差を最大にするのが最適である、という観点から解析したものです。両者の違いについては一番下で少し議論することにします。
ここまで述べてきたように、光合成能力(ここでいう光合成能力とは、大気条件下、光飽和下の光合成速度)が高ければ適応的である、という単純なものではありません。光合成能力の高低の決定において重要なコストと考えられているのは窒素です。高い光合成能力には相応の光合成系タンパク質が必要です。タンパク質を作るためには窒素が必要となります。光合成系のタンパク質の窒素は葉の窒素の実に約半分を占め、光合成能力と窒素含量の関係は高い相関を示します。また、生態系においては、窒素は最も不足しがちな元素の一つです。「窒素をいかに効率よく利用するか」が植物の適応度を決める一因である、というのは広く認識されていることだと思います(Field & Mooney 1986)。ここでは、葉にどれだけ窒素を投資するべきか?ということを考えてみましょう。
Mooney & Gulmon(1979)は「植物は異なる環境で光合成能力をいかに変えるべきか」という経済学的観点から解釈を試みました。葉の窒素含量を増せば高い光合成能力を実現できますが、高い光合成能力を持っていても実際の光合成速度を高くすることができるとは限りません。例えば弱光条件では、光合成速度は光強度に律速されますので、高い光合成能力を持っていてもそれが必ずしも炭素の獲得に結びつきません。したがって、コストを抑える、という意味で窒素含量や光合成能力が低いほうが適応的です。一方強光条件では、光合成能力を高めれば相応の光合成速度の増加が見込めますので、高い窒素含量が有利です。実際に、強光条件下の葉ほど窒素含量が高いことが知られています(DeJong & Doyle 1985, Hirose & Werger 1987a, Hikosaka et al. 1994)。
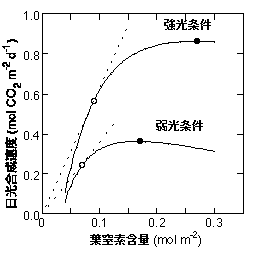
生育環境によって窒素含量がどのように調節されるべきかをより詳細に検討してみましょう。葉の光合成能力と葉面積あたりの窒素含量が高い相関を示すことから、窒素含量と一日の光合成量の関係の推定が可能です(Hirose & Werber 1987a)。下の図はホウレンソウの葉のデータ(Terashima & Evans 1988)をもとにしたモデルによる一日の光合成量と窒素含量の関係です(Hikosaka & Terashima 1995)。窒素含量が高い葉ほど一日の光合成量が多いのですが、一日の光合成量は窒素含量に対し曲線となり、最大値(黒丸)を持ちます。高窒素含量で光合成量が下がってしまうのは呼吸速度が窒素含量とともに増加することを仮定しているためです。光合成量を最大にする窒素含量は弱光条件ほど低いです。これは、先ほど述べたように光合成能力が高くても光強度に光合成速度が律速されることが多いからです。
まず、一つの光環境に絞って見てみましょう。この図の白丸は曲線に対して減点を通る接線(点線)が交わる点であり,窒素あたりの光合成量が最大になるポイントです(Hirose 1984)。このポイントより窒素含量が高くても低くても光合成における窒素の利用効率は悪くなります。このことを理解するために、1gの窒素を持っている,ある葉を仮定しましょう。この葉は戦略として葉面積を変えることができます。しかし、葉面積を広げると当然面積あたりの窒素含量は下がってしまうので、葉面積と面積あたりの窒素含量の間にはトレードオフが成り立ちます。白丸よりも窒素含量が2倍大きい葉を考えてみましょう。この葉は葉面積あたりの光合成量は高いものの、2倍には達せず、葉面積が半分であるために結局葉あたり(窒素1gあたり)の光合成量は小さいです。白丸よりも窒素含量が低い場合も、葉面積の拡大が面積あたりの光合成量を保証できず、やはり葉あたりの光合成量は小さくなります。したがって「窒素あたりの光合成量を最大にする」ことが適応度を最大にすることと同義であれば、ある一つの光条件に対し一つの最適な窒素含量しかとりえないことになります。この窒素含量が最適な窒素含量である、ということになります。
次に、異なる光条件の葉を比べてみましょう。黒丸の窒素含量を比べると、強光条件の方が高いことがわかります。したがって、富栄養条件では窒素含量は強光条件のほうが窒素含量が高くなることが予測されます。一方、白丸の窒素含量を比べると、強光条件のほうが高いですが、あまり差はありません。このことから、貧栄養条件では窒素含量はあまり光条件に依存しないことが予測されます。これはホウレンソウにおける実験結果(Terashima & Evans 1988)と一致します(Hikosaka & Terashima 1995)。
以上の話は彦坂(1997)にも書きましたのでそちらもどうぞ。
ここでは、Cowan (1997) に基づいて最適な気孔開閉調節を考えてみましょう。気孔と光合成・蒸散の関係についてはこちらをご覧ください。
気孔を開けることにおいては、光合成による炭素獲得がベネフィットで、蒸散による水の損失をコストと考えることができます。そこで「ベネフィット-コスト」を最大化するように気孔開閉を調節することが最適であると考えられますが、炭素と水蒸気では引き算はできません。そこで、水を吸収するために必要なコストを炭素に換算するためのパラメータλを導入します。λは水を吸収するための根の構成コストや維持コスト、その他輸送などにまつわるあらゆる炭素コストを、蒸散される水当たりで表した値です。
気孔コンダクタンスと光合成・蒸散の関係は、図にすると下のようになります。
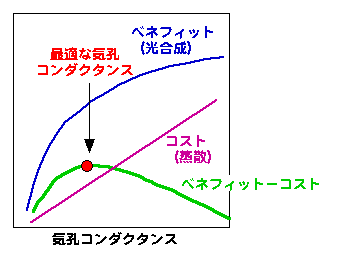
蒸散速度は気孔コンダクタンスに比例しますが、気孔コンダクタンスが無限大になっても葉の中のCO2濃度は外より高くなることはないため光合成速度と気孔コンダクタンスの関係はアタマ打ちになります。そうすると光合成速度と蒸散にまつわるコストの差が最大になるところが最適な気孔コンダクタンスということになります。
最適な気孔コンダクタンスは以下のようにして得ることもできます。
A-λE = 最大
ここでAは光合成速度、Eは蒸散速度です。λは定数ですが、環境条件によって変化すると考えられます。例えば、乾燥条件では根あたり吸収できる水の量は少ないので、λは大きくなります。式1を満たす解はA-λEの極大を求めることによって得られるますので、A-λEの微分が0になる条件を求めます。解は以下のようになります。
λ= dA/dE
ここで、dA/dEは縦軸にA、横軸にEをとったときの曲線の傾きです。つまり、λが一定である限り、気孔の開閉はdA/dEが一定になるように調節するのが最適だということになります。光合成速度と蒸散速度の関係は以下のような図になります。
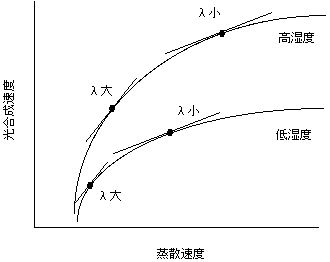
上で説明したように、蒸散と光合成の関係は頭打ちになります。さらに、光合成と蒸散の関係は湿度に依存します。光合成と気孔開度の関係は湿度の影響を受けませんが、湿度が下がると、同じ気孔開度でも蒸散速度が高くなります。このため、湿度が下がると同じ光合成速度でも蒸散速度が高くなるわけです。
日中湿度が変化するような条件では気孔開度をどう調節すればいいか考えてみましょう。相対湿度が下がると、気孔を開けたままではdA/dEが下がってしまいます。そこで気孔を閉じ気味にして蒸散速度を下げ、dA/dEを上げなければいけません。昼寝現象は以上のように説明できます。
また、乾燥条件に生育する植物と、湿潤条件に生育する植物を考えてみましょう。上述のように、両条件では水を吸収するためのコストが異なり、乾燥条件のほうがlが高いと考えられます。葉の周囲の環境が同一条件だったとしても、乾燥条件の植物のほうがlが高いので、気孔を閉じ気味にしdA/dEを高く保つ必要があることが予測されます。
このモデルによる予測は実際の気孔開度の調節をよく説明しています(Cowan 1986)。ただし、植物がdA/dEをどのように感知しているかはたぶんまだわかっていないんだと思います。なお、Cowanの一連の研究では、λのかわりに1/λを使っていることを申し添えておきます。
「窒素利用効率」は様々に定義されてきました。私自身もこれまでに三種類ほど使ってきています。現在普及している「窒素利用効率」は二種類です。一つは、葉の含有窒素あたりの最大光合成速度です(葉面積あたりの飽和光合成速度を葉の窒素含量で割る)。光合成窒素利用効率 Photosynthetic nitrogen use efficiency とし、PNUEと略されます。PNUEは、光合成において窒素が光合成系タンパク質を作るために必要が元素であることと窒素含量と光合成能力の間に高い相関を持つことを念頭において使われています。PNUEについてはこちらに詳しく書きましたので、ここでは省略します。
もう一つの定義は、個体(あるいは個体群)が吸収した(あるいは放出した)窒素量あたりの成長量です。個体レベルの現象を扱う場合、この定義が妥当であると考えられています。この項でも、以下で扱う窒素利用効率はこの定義を使うことにします。
窒素利用効率を最初に定義したのはHirose (1975) のようです。以下は余談になります。広瀬さんはうちの研究室のボスでしたが、私は窒素利用効率を最初に定義したのが広瀬さんであることを知りませんでした。私は以前オオオナモミ群落で窒素利用効率を測定したことがあり、広瀬さんに原稿を持っていったところ、「これを読んでみて下さい」と言って渡されたのがHirose (1971) とHirose (1975) でした。前者は広瀬さんの修論を投稿論文にしたもの、後者はD論です。残念ながら国際誌に投稿されたわけではなかったため、あまり広くは知られていませんが。とはいえ、Vitousek やBerendse, Aertsといった専門家達には最初の仕事として認知されているようです。
普及の最初のきっかけはChapin (1980) です。一年草では窒素のロスがそれほど多くないので、植物が持っている窒素の量=窒素の吸収量とみなすことができます。また、一年草のバイオマスはそれまでの成長量とみなすことができますので、個体のバイオマス/窒素比、つまり個体の窒素濃度の逆数を窒素利用効率としました。さらに、Vitousek (1982) の研究があります。彼は森林レベルの窒素利用効率を調べたのですが、彼が窒素利用効率のかわりに使ったのはリターの窒素濃度の逆数でした。ある程度安定な森林では、植物が吸収する窒素の量と植物がリターなどで放出する窒素の量はほぼ同様の値を持つと考えられます。同様に植物の成長量とリター放出量が等しいと仮定すると、リターの量/リターの窒素量比は窒素利用効率に近い値を出すと考えられます。窒素の放出量は比較的測定しやすいので、この手法は多くの研究で使われることになります。
窒素利用効率の概念を生態学的に非常に面白いものにしたのがBerendse and Aerts (1987) です。それは次に紹介することにしましょう。
Berendse and Aerts (1987) の窒素利用効率の定義も、基本的には「吸収(放出)窒素量あたりの成長量」になります。彼らはさらに窒素利用効率を二つのパラメータの積で表します。
NUE = dW/dN = (1/N × dW/dt) × N dt/dN
dはデルタと考えたほうがいいでしょう。ぱっと見でよくわからない式ですが、まず、Nとdtを消去するとdW/dtになることを確認して下さい。右辺のカッコ内のパラメータは、成長速度(dw/dt)を個体が持つ窒素量(N)で割ったものです。つまり窒素あたりの成長速度ということになります。これはかつてIngestad (1979) によって窒素生産力(Nitrogen productivity)と定義されたものです。その右のパラメータは、個体が持つ窒素量を窒素吸収(あるいは放出)速度(dN/dt)で割ったものです。このパラメータが意味するのは、ある窒素原子が植物体に吸収されてから、体外に放出されるまでの平均時間を表します。平均窒素滞留時間(Mean residence time of nitrogen)と呼ばれます。平均窒素滞留時間の逆数がターンオーバー速度になります。窒素生産力をNP、平均窒素滞留時間をMRTと略し、NUE = NP × MRT というように表します。
この式は、いわゆる成長解析のようにNUEを分解しているわけですが、その分解の仕方が非常に秀逸であるため、以降の研究に大きな影響を与えました。秀逸な点は、この分解が植物の窒素利用に二つの全く異なる生物学的側面があることを簡単に示したからです。一つは、植物が持つ窒素の成長への貢献です。このHP内のあちこちで述べているように、葉の窒素は光合成生産に直結します。植物が持つ窒素を多く光合成系に投入すればするほど高い光合成生産ができ、高い窒素利用効率を実現するはずです。一方で、窒素利用効率を高めるには別の方法があります。それは、吸収した窒素をできるだけ放出しない、つまり、長時間体内にとどめておけば(たとえ光合成生産がよくなくとも)、窒素利用効率は高いと考えられます。Berendse and Aertsの式は、窒素生産力が前者を、平均窒素滞在時間が後者を表します。
さらに、Berendse and Aerts (1987) では、窒素生産力と平均窒素滞在時間の間にトレードオフがあることを指摘しています。つまり、一つの植物では、片方を高めるような戦略を採ると、もう片方が下がってしまう、ということです。彼らはオランダの異なる栄養条件に成立する草原で数種の植物のNUE、NP、MRTを測定し、測定した植物の間ではNUEがそれほど違わないこと、NPが高い種はMRTが低いことを示しました(ただし、NPとMRTの間のトレードオフは観察されたりされなかったりで、本当にそういう関係があるのかは、実験結果からはまだ確立されていません。Garnier and Aronson 1998)。
窒素生産力が高い植物は、いわゆるfast-growing species(理想的な条件で育成したとき、相対成長速度が高い種)にあたるようです(Poorter et al. 1990)。攪乱が起こったあとに、素早く侵入し、高い成長速度で多種を圧倒する、というのがこういった種の生態学的性質です。窒素生産力を高めるには二つの方法があります(Hirose 1988)。一つは、個体が持つ窒素の多くを葉に投資することです。葉の窒素が多いほど多く光合成ができます。もう一つは、窒素あたりの光合成能力(PNUE)を高めることです。PNUEについてはこちらに記しましたのでご覧下さい。Poorter et al. (1990) やGarnier et al. (1995) の研究によって、窒素生産力が高い植物はPNUEも葉への窒素投資比も高い傾向があることが示されています。また、「光合成能力のコストとベネフィット」で紹介した窒素あたりの光合成も窒素生産力に関連していると考えられます。光合成の窒素利用は個体レベルの窒素利用効率を考える上では、パラメータの一つにすぎず、光合成の窒素利用が高いイコール個体窒素利用効率が高い、というわけではないことに注意してください。
一方、平均窒素滞在時間を長くするにも二つの方法があります。一つは、枯死器官からの窒素回収の効率を高めることです。植物は器官が枯死するときに、そこから窒素やリンなどの栄養塩を回収します。持っていた窒素のうち、枯死時までに回収された窒素の割合を、窒素回収効率といいます。回収効率が高ければ、体内から出ていく窒素の量が少なくなり、窒素滞在時間が長くなります。もう一つの方法は、葉の寿命を長くすることです。いくら枯死時に窒素を回収するとはいえ、全ての窒素を回収できるわけではありません。葉ができてから枯死するまでの時間が長ければ(=葉の寿命が長ければ)、平均滞在時間が長くなることになります。Escudero et al. (1992) は地中海性気候に生育する樹木数十種について平均窒素滞在時間・窒素回収効率・葉の寿命を調べました。窒素回収効率は種によって様々な値を持ちますが、平均窒素滞在時間とは相関がなく、窒素滞在時間と強く結びつくわけではないようです。一方、葉の寿命と平均窒素滞在時間の相関は強く、葉の寿命が長い植物で平均窒素滞在時間が長いことが示されています。
このような違いは、Reichらによる光合成の種間差の話と非常に近いと思われます。Reichらは葉の寿命が長い種では光合成能力やPNUEが低いことを報告しています。葉の寿命が直接平均窒素滞在時間を、PNUEと窒素生産力の間に相関があるのなら、平均窒素滞在時間と窒素生産力の間に負の関係(つまりトレードオフ)があることはほぼ間違いないと思われます。前項では「本当にそういう関係(トレードオフ)があるのかは、実験結果からはまだ確立されていません」と書きましたが、これは単に測定例が少ないから確立されていないだけだと私は考えています。
以上のように、窒素利用効率というのはいろいろな性質を反映していて、なかなか面白いパラメータですが、このパラメータを最大化することにどういう意味があるのかを説明することは簡単ではありません。現在の進化生物学では、適応度(子孫の数)を最大にする性質が進化の過程で選択されたと考えます。窒素利用効率を最大化することと適応度最大化がイコールかどうかは真剣に議論されたことがありません。しかし、窒素利用効率、特に平均窒素滞留時間が重要であることを示唆する材料はいくつかあります。
一つは、成長速度の高低では貧栄養条件への適応をはかれないことです。我々は、分布域から、ある種が貧栄養条件に適応しているのか、富栄養条件に適応しているのかを経験的に判断することができます。ドイツのような植物社会学が発達した国では、N-numberといって、植物がどのような栄養条件に適応しているのかを10段階評価で表した人がいます(Ellenberg 1979。原著はドイツ語で、私も読んでいません)。このN-numberと生理生態学的なパラメータは高い相関があります。例えば、最適な環境で育成したときの相対成長速度とN-number(高いほど富栄養適応)の間には高い正の相関があります(Poorter and Remkes 1990)。このことは、生理生態的な性質がニッチの決定に強く影響していることを示唆します。N-numberが高い植物は、好条件下での相対成長速度が高く、富栄養条件で競争に強いだろうということは一目瞭然です。では、貧栄養条件に適応している植物はどうでしょうか? 単純には、貧栄養条件に適応している植物は、富栄養に適応している植物に比べ、貧栄養条件での相対成長速度が高いことを期待します。しかし、必ずしもそうではないようです(van der Werf et al. 1993)。単なる成長の高低ではニッチは説明できないわけです。
もう一つは、平均窒素滞留時間の重要性です。Aerts & van der Peijl (1993) は、数理モデルを使って窒素利用の違いが植物の成長にどのような影響を与えるかを考察しました。Aerts (1990) のデータを使い、窒素利用効率が等しい二種を考えます。ただし、片方は窒素生産力が高く、もう片方は平均窒素滞留時間が長いとします。貧栄養条件を仮定し、両種のバイオマスが時間とともにどのように変化するかを予測しています。その結果は、窒素生産力が高い種は急速に成長するが、すぐにバイオマスが頭打ちになってしまいます。一方、平均窒素滞留時間が長い種は、初期成長は遅いものの、あとで群落のバイオマスが高くなります(どうしてこうなるかは、ちょっと今はわかりません。後で調べます)。したがって、長期的に見ると平均窒素滞留時間が長いことが有利である可能性があります。
Aerts and Chapin (2000) は、窒素利用効率だけでも異なる栄養条件への適応を考えることはできない、としています。Aerts (1990) では栄養条件が異なる場所に生育する植物が似たような窒素利用効率を持ち、窒素生産力と平均窒素滞留時間の間にトレードオフがあったため、このような主張をしています。ただ、貧栄養条件に生育する植物で窒素利用効率が高い傾向があることもあるので(Vitousek 1982)、窒素利用効率そのものにも意味があると考えるべきかもしれません。いずれにせよ、窒素利用効率・窒素生産力・平均窒素滞留時間の三者を考えることで様々な栄養条件への適応を考えることができるようになるだろう、という雰囲気は充分あります。
水利用効率も様々に定義されていますが、そのバリエーションは窒素利用効率ほどではありません(たぶん)。農学では、乾物生産量(あるいは農産物収量)を使われた水の量で割ったものを水利用効率として使用しているようですが、あいにく私はこの使用例を知りません(農学には知識がないもので)。特に乾燥地では水の利用は切実な問題であることと、農作物は与えた水の量を定量化しやすいのでよく使用されるものと思います。しかし自然生態系を扱う場合は植物が利用した水の量を定義するのは難しいでしょうねえ。ただ、近年は樹木の幹の中で水が吸い上げられる量を測ることができるらしい(サップフローメータ)ので、不可能ではないでしょう。
葉レベルでは、光合成速度を蒸散速度で割ったものを水利用効率と定義します。失われる水の量をコスト、得られる光合成量をベネフィットと考えるわけです。本項では葉レベルの水利用効率を主に扱うことにします。
まずは、水利用効率がどのような要因に支配されているかを考えてみましょう。まずこちらを読んで、水と二酸化炭素の拡散に関する基本を抑えておいて下さい。光合成速度Pは以下の式で表されます。
P = gs(Ca - Ci)
ここで、gsが気孔コンダクタンス、Caは葉の外の二酸化炭素濃度、Ciは葉内細胞間隙の二酸化炭素濃度です。さらに、蒸散速度Eは以下の式で表されます。
E = gh(Hi - Ha)
ここで、Hiが葉の中の水蒸気密度、Haが葉の外の水蒸気密度です。gsとghの間には以下の関係があります。
1.6gs = gh
したがって水利用効率WUEは以下の式で表されることになります。
WUE = P/E = (Ca - Ci)/(Hi - Ha)/1.6
右辺からは見かけ上光合成速度もコンダクタンスもなくなってしまうので、ずいぶん単純化されたイメージを受けます。が、WUEは様々な要因の影響を、複雑に受けます。以下は、直接影響を与えるものを書きます。
1)大気湿度
葉の中では、水蒸気密度はほぼ飽和していると考えられています。葉温と気温が等しい場合、Hi-Haは大気飽差(VPD。分圧で表しますが、密度と同じように考えて下さい)に相当します。飽差が大きいということは、空気が乾いているということになります。もしCiが変わらなければ、空気が乾き、飽差が大きいほど蒸散が大きくなり、水利用効率が下がることになります。水利用効率は高温で下がることが多いようですが、これは主に高温で飽差が大きくなりやすいことによるのだと思います。
2)気孔コンダクタンス
気孔コンダクタンスは光合成速度と蒸散速度の両方に影響を与え、両者とも気孔コンダクタンスが下がると低くなります。水利用効率はどうなるかというと、右辺を見て下さい。気孔コンダクタンスはHi-Haにはそれほど影響しません(厳密には、蒸散で葉温を下げることによりHiを下げる作用があります)、Ciに大きく影響します。気孔が閉じるとCiが下がるため、式の形からすると、気孔コンダクタンスが下がるほど水利用効率が高いことになります。直観的な理解としては、気孔コンダクタンスが低下すると蒸散速度が比例して低下するのに対し、光合成速度の低下はそれほど大きくない(逆にいうと、気孔コンダクタンスが二倍になったからといって光合成速度が二倍になるわけではない)ことによります。
3)光合成速度
理論上は、光合成速度が低くなる場合は水利用効率は低くなるはずです。ただ、光合成速度が低下するときは、多くの場合気孔コンダクタンスも低下するので一概に言えるわけではありません。高CO2条件では水利用効率は一般に上がります。これは、高CO2濃度では光合成速度が高いことと、気孔コンダクタンスが低下して蒸散速度が低下することの両方によります。飽差が一定で、水ストレスが起きていない場合、気孔コンダクタンスはCi/Ca比を一定に保つように変化する傾向があります。Ci/Ca比が一定ならば、Caが大きいときほどCa-Ciは大きくなります。
炭素安定同位体13Cの含有量は、測定機器を持っている研究室なら比較的容易に測定できます。そんなわけで13Cの含有量からいろいろな情報を得ようとする研究者は多くいます。安定同位体の利用についてはこちらに詳しく書いています。光合成に関しては、葉に含まれる13Cの含量を示すδ13Cを水利用効率の指標に使うことがあります(ただし、C3植物のみに適用可能)。 なぜδ13Cから水利用効率を推定できるかというと、このパラメータとCiの間には強い相関があるためです。理論的な数式展開はこちらに書きましたのでご覧下さい。実測したCiと13Cから推定したCiの間に相関があることもいくつかの研究で示されています(Farquhar et al. 1989)。
安定同位体を使うことには、測定が容易ということの他にもう一つメリットがあります。それは、安定同位対比が示す水利用効率は、瞬間値ではなく積算値だということです。光合成測定によって得られる水利用効率はしょせん瞬間値です。一日の間には湿度・気温・光強度に大きな変化があり、ある時間での水利用効率が全体を反映しているとは限りません。その点、葉の13C含量はそれまでに植物が炭素を吸収するときのCiの平均値を反映していると考えられます。ただ、厳密に考えると、葉を構成する炭素の大部分はその葉が吸収した炭素ではなく、その葉が生まれる以前に別の葉が同化したものですから、長期的な環境変動が大きい場合には逆に何を見ているのかわからなくなりかねません。ある葉の13C含量がいつの環境のCiを反映しているかを研究している方もいるくらいです。
13Cの使い方で気をつけなくてはいけないことは、13Cが語るものはあくまでCiであり、水利用効率ではない、ということです。葉の13Cを測っただけではHaもHiもわかりません。同一環境に生育している植物でならHa-Hiは等しいと考えていいだろう、ということでCiと水利用効率の関係は1:1の関係にあるとみなせるわけです。しかし、異なる環境に生育する植物を比較する場合は、δ13Cが水利用効率を示すとは限らないことになります。まあ、Ciが低いということはそれ自体が気孔が閉じ気味であることを示すので、それに基づいた考察をするのは問題ないと思いますけど。
私は水利用については専門ではない、ということを認識しつつ以下をお読み下さい。
資源利用効率の考え方に基づけば、水資源が不足している場所を好む植物はWUEが高い、という発想が自然です。実際そう考えていた先達がいるようですし、私も漠然とそう思っていました。ところが現実は必ずしもそうではないようです。私の手元にあるのはDeLucia and Schlesinger (1991) という論文です。この論文のイントロダクションは、学生の頃の私には納得いかないことばかりでしたが、水利用について少しづつ知識が増えた今ではなんとなく「そうなのかな」と思うようになりました。この論文が提示する仮説は、上と逆で、「乾燥耐性がある植物ほどWUEが低い」です。根拠はいくつか挙げられています。一つは、PNUEとWUEのトレードオフです。PNUEとは、上に書いたように葉の光合成速度を窒素含量で割ったものです。窒素含量は気孔開閉の影響を受けないので、気孔を閉じるとPNUEは下がることになります。つまりWUEを上げることはPNUEを下げるという不利を伴うことになります。もう一つは、水を奪い合う競争です。WUEを下げるということは多くの場合蒸散の低下→吸水量の低下を伴います。隣接個体と水を奪い合う競争をしている場合、自分が吸水量を低下させると隣接個体の吸水量を相対的に上げることになり、不利になります。実際8種ほど比較してみると、乾燥耐性とWUEの間には負の相関がありました。
彼らは具体的に言及していませんが、水ポテンシャルの低下に対する適応も重要かもしれません。葉からの蒸散に根からの吸水が追いつかない場合、水が過剰に出ていくことによって水ポテンシャルが低下します。種によっては水ポテンシャルが低下するとすぐに光合成能力が下がってしまうものと、比較的低いポテンシャルでもある程度の光合成速度を維持できる植物もいます。葉の水ポテンシャルが低いほうが潜在的な吸水能力が増すということもあるので(このあたりの詳しいことは、私の知識が充分になったら書くことにしましょう)、かえってそのほうが有利なのかもしれません。ただし、低い水ポテンシャルで光合成を維持するためには何らかのコストがかかるはずです。
以上のように、資源利用効率が必ずしも適応の度合いを示すとは限らない、ということでご理解下さい。現在どのようなコンセンサスが得られているのかは知りません。
上で述べたように、コストとベネフィットの観点から資源利用の効率をいかに評価するかについては二つの考え方があります。一つは、ベネフィットとコストの差を最大にする戦略が最適であるという考え方です。もう一つが、ベネフィットとコストの比を最大にする戦略が最適であるという考え方です。窒素利用効率や水利用効率など、資源利用効率は後者にあたります。ここでは両者の違いを少し考察してみましょう。
まず両者を図を使って比較してみましょう。コストをX、ベネフィットをYとする線を描いてみます。一般に、コストとベネフィットの関係は頭打ちの曲線になると考えられます(左下図)。コストの過大投入は無駄を引き起こしやすいためです。このとき、ベネフィット/コスト比を最大にするのは、原点から引いた直線が曲線に接する点になります。一方、ベネフィットとコストを同じ単位で比較できると仮定すると、コストはY=Xのベネフィットとコストの差を最大にする点は右下図の赤線になります。ベネフィット-コスト差を最大にする点は必ずベネフィット/コスト比を最大にする点よりも高くなります(低くなるときはベネフィット-コスト差は負の値)。
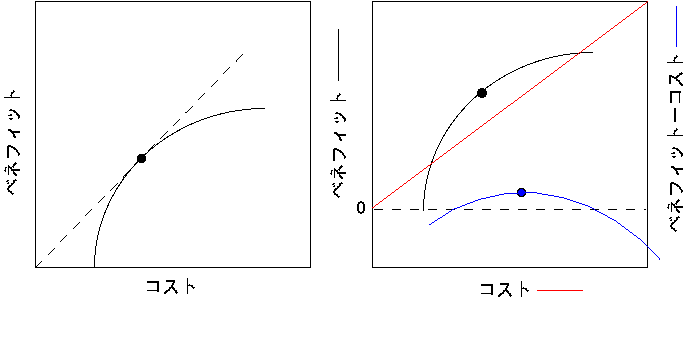
論理的にいえば、適応度を考える上で妥当なのはベネフィット-コスト差であると考えられます。例えば、ベネフィットを光合成、コストを呼吸と考えるような単純な系ならば、植物が実際に示すパフォーマンスである成長量はベネフィット-コスト差となります。適応度が成長量に比例するならば、ベネフィット-コスト差が最大になる条件が有利ということになります。
しかしベネフィット-コスト差の適用には大きな問題があります。それは、ベネフィットとコストを同じ単位で表すことが困難であることが多いことです。上で述べた蒸散と光合成の例でいうなら、二酸化炭素の量から水蒸気の量を引くことはできません(ちなみに、モルでいうと、後者は前者の100倍くらい大きいです)。Cowanのモデルでは、蒸散量を炭素量に置き換えるために、λというパラメータを採用しました。蒸散をするためには、根へのバイオマスの投資、維持のためのコストなどが必要であると考えるわけです。それによって光合成と蒸散量とを同じ単位で比較できるようにしました。Cowanのモデルが成功したのは、λの値があいまいでも最適解を(なんとなく)理解できるからですが、「この環境でこのように気孔を開けるのがよい」、ということを定量的に予測するためにはλの値を知る必要があります(Cowanによるモデルの検証では、当てはまりそうなλをあらかじめ仮定しています)。現実にはλを推測するのは困難に思えます。
このようにベネフィット-コスト差を定量化するのが困難な場合に、ベネフィット/コスト比を代用する場合があると考えることができます。資源が非常に乏しいとき、ベネフィット-コスト差は小さくなると考えられます(例えば、水分が不足している場合は根を多く作らなくてはいけないため、コストがかかる)。ベネフィット-コスト差が0に近づけば近づくほどベネフィット-コスト差とベネフィット/コスト比を最大にする解は近くなります。
これとは別に、ベネフィット/コスト比を最大にする解が適応度の指標としてふさわしい場合もあります。それは、うまく説明できないのですが、例を挙げると以下のような場合です。葉面積あたりの窒素含量をコスト、葉面積あたりの光合成量をベネフィットとした場合、窒素コストを炭素量に換算可能であるとしましょう。個葉レベルではベネフィット-コスト差を最大にする窒素含量と、ベネフィット/コスト比を最大にする窒素含量は異なります。上の議論ではベネフィット-コスト差を最大にすることが適応度最大に近いように思いますが、実際は必ずしもそうではありません。話を簡単にするために、炭素換算した窒素コストがゼロに近いと考えてみましょう。そうすると、ベネフィット-コスト差を最大にする窒素含量はかなり大きな値になります。しかし、ここでの光合成量はあくまでも葉面積あたりであることに注意して下さい。葉が持っている窒素量を一定とし、個体は「葉面積が小さいが窒素濃度が高い葉」あるいは「葉面積が大きいが窒素濃度が高い葉」を作ることができると考えましょう。このとき、高い窒素量は、葉面積あたりでは高い光合成量を持っていますが、葉全体で比べると、ベネフィット/コスト比を最大にする窒素含量が最も高い光合成量を持ちます(詳しい説明はこちら)。つまりこの場合、ベネフィットとコストを葉面積あたりで表しているため、「葉面積あたりの」ベネフィット-コスト差を最大化することが「個体の」ベネフィット-コスト差を最大化することと一致していないのです。このような状況は、葉に投資した窒素量と光合成量の関係が一定ではない(同じ窒素量が投資されていても、その窒素をどのようにふりわけるかによって光合成量が変わってしまう)場合に起こります。これは典型的なトレードオフの関係で、しばしばパイのトレードオフと呼ばれます。パイの一切れを大きくすると数が少なくなり、数を増やすと一切れの大きさが小さくなります。
このように、ベネフィットとコストの関係は状況によってその意味が変わります。安易に資源利用効率を定義することは危険なことで、それが適応度とどのような関係にあるかは慎重な考慮が必要です。しかし、上に書いたように、生理生態が扱う分野ではベネフィットとコストを同じ単位で換算することは困難な場合が多いですし、材料として栄養成長期の個体を扱う場合は、適応度との関係も明らかにできないこともほとんどです。このような場合に資源利用効率はある一つの目安として使うべきものと私は考えています。