光合成の生理生態学講座
気孔コンダクタンスの物理学(200602 12)
光合成に対する気孔コンダクタンスの影響(200602 12)
気孔の環境生理学(200602 12)
なぜ気孔開度の不均一性を考えなくてはいけないのか(200102 21)
「不均一な光合成」の発見(200103 02・200104 07改変)
homobaric leaf と heterobaric leaf(200103 12)
「不均一な光合成」のその後(200103 20・200104 07)
光合成をするには基質である二酸化炭素が必要になります。二酸化炭素は、空気中から葉緑体まで受動的、つまり濃度勾配に依存して拡散していきます。さかんに光合成をしている葉では、葉緑体のCO2濃度は大気中のCO2濃度(350-370 ppm)の約半分近くまで落ちています。つまり、それだけの抵抗がかかっているわけです。「二酸化炭素と光合成」で説明したように、CO2濃度の低下は光合成の効率を下げます。なぜ光合成を低下させてまでCO2の拡散を妨げているのか?という疑問を持ちつつ本項を読んでいただければ幸いです。
まず、ここでCO2が葉の中を拡散する経路について述べておきます。一般的に、葉の表皮細胞はCO2をほとんど通しません。CO2はほとんど全て気孔を通って葉の中に入ります。葉の中は細胞がぎっしりとつまっているわけではなく、細胞と細胞の間は気相の空間があります。これを細胞間隙と呼びます。気孔を通ったCO2は、細胞間隙を通って葉肉細胞の細胞壁にたどりつきます。このあと、細胞壁を通って、細胞膜を通過し、細胞質に入り、葉緑体包膜を通過してストロマに達します。
このページでは主に気孔の影響について記します。大気から葉内細胞間隙までのCO2拡散にまつわる話です。
まずは「拡散とは何か」という話から始めましょう。ただ、私は高校の理科I以来物理を学んだことがありません。こーゆー人間が物理の解説をしていいのだろうか、という問題があるので、眉に唾しながら読んで下さい。とはいえそれほど難しいことは書きません(書けないから)。
分子というのは常に運動しています。ある空間内に分子の濃度の不均一性がある場合、分子の移動が起こります。これが拡散です。一次元の移動を仮定したとき、ある物質が距離xで離れている二点間を移動する速度Jは以下の式で表されます。
J = -D (C2 - C1)/x
ここで、C1、C2はそれぞれ二点での物質の濃度、Dは拡散係数です。なおこの式はFickの第一法則と呼ばれます(本物は、濃度差をxで偏微分していますが)。Dが大きいほど「拡散しやすい」といいます。光合成の場合は、「ある物質」はCO2で、Jが光合成速度そのものになります。ところで、-D/xをひとまとめにすると、これは速度と濃度差の比例定数となります。これがコンダクタンスです。書くまでもありませんが、
J = g(C2 - C1)
となります。コンダクタンスの逆数が抵抗です。これは電気でいうオ-ムの法則に相当します。電気の場合は、Jが電流、C2-C1が電圧、1/gが電気抵抗、というわけです。CO2の拡散も、オ-ムの法則のアナロジーとして考えることができます。
気孔は、葉肉細胞と外界をつないだり遮蔽したりする役割を持ちます。下に書くように、条件によっては気孔は完全に閉じてしまい、ガスの交換が起こらなくなります。気孔の開閉がガスの拡散に表す影響を、上に述べたコンダクタンスを使った式で表すわけです(Gaastra 1959)。
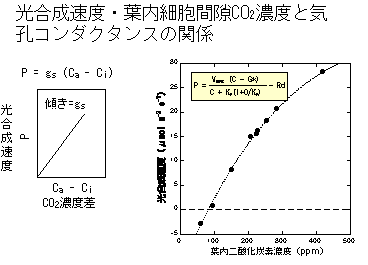
P = gs(Ca - Ci)
ここで、Pが光合成速度、gsが気孔コンダクタンス、Caは葉の外のCO2濃度、Ciは葉内細胞間隙のCO2濃度です。
さて、Ciやgsはよく計測されるパラメータです。特に、最近は自動で計算してくれる携帯光合成測定装置が出回ってますので、どういう理屈でこれらのパラメータが計算されているか理解しないで使っている人も多いのではないかと推測されます。PとCaはCO2濃度を測定すれば計算できますが(葉をチャンバーの中に入れて、チャンバーから出てきた空気がCaで、チャンバーに入る空気のCO2濃度と出てくる空気のCO2濃度の差が光合成速度)、gsやCiはCO2濃度を測定するだけでは計算できません。gsは水蒸気の出入りを調べることによって求めます。
CO2に対するコンダクタンス(gs)と水蒸気に対するコンダクタンスghは比例関係にあります(このときのコンダクタンスはmol m-2 s-1という単位で表されます)。
1.6gs = gh
この式の意味するところは、水のほうがCO2より1.6倍気孔を通過しやすいということです。この違いは単純に分子の大きさの違いと考えてよいと思います。水蒸気の拡散も同様な式で表されます。ただし、水蒸気は葉の中から外に出ていくという点が異なりますが。
E = gh(Hi - Ha)
ここで、Eが蒸散速度、Hiが葉の中の水蒸気密度、Haが葉の外の水蒸気密度です。EとHaは、水蒸気密度を直接測定することで求めることができます。ここまではCO2の場合と同じです。水蒸気の場合、CO2の場合と違う点は、葉の中の水蒸気密度を推定する方法があることです。ここで、中学校か高校で習ったはずの、「飽和水蒸気密度」というのを思い出して下さい。飽和水蒸気密度とはその条件において水蒸気が大気中で存在可能な最大の濃度です。水蒸気の供給が飽和水蒸気密度を上回れば、存在できない水蒸気は水となります(結露)。飽和水蒸気密度は気温と気圧に依存し、高温ほど高くなります。葉内では、水蒸気は飽和していると考えられています。葉っぱの温度を測定し、その温度での飽和水蒸気密度を計算すれば、それが葉の中の水蒸気密度(Hi)ということになります。Hiがわかれば、ghがわかり、gsがわかり、Ciがわかる、というわけです。つまり、Ciを計算するためには、CO2濃度、水蒸気密度、葉温の測定を正確にしなくてはいけないわけです。
この方法で本当にCiが計算できているのか、というのは非常に賢い方法で証明がなされています。両面に気孔がある葉っぱを使い、片面を「閉鎖系」に、もう片面を「開放系」にします。どういうことかというと、「閉鎖系」とは葉っぱにおわんをかぶせたと思って下さい。外界との連絡はありません。「開放系」とは、おわんをかぶせるのですが、そのおわんには空気の入り口と出口があります。こちらは通常の光合成を測るときと同じだと考えて下さい。「閉鎖系」のほうは、放っておくと、平衡に達して、葉の中と同じ状態になるはずです。「開放系」を使って上の式を使ってCiを測定します。これは「閉鎖系」のおわんの中のCO2濃度と同じになる、ということで理論が検証できるわけです(この話は昔誰かのセミナで聞いただけで、文献を思い出せません)。
「二酸化炭素と光合成」ではCO2濃度が変わると光合成速度がどのように変わるのかについて解説しました。一方、こちらでもP = gs(Ca - Ci) というCO2濃度と光合成速度の両方が出ている式を出しました。二つの式はどういう関係にあるのでしょうか。わかりやすいかどうかよくわかりませんが、講義で使っている説明をそのまましてみましょう。
P = gs(Ca - Ci) というのを言葉で表すと、「葉内外のCO2濃度差と光合成速度は比例する」となります。比例定数がgsなわけです。この式は拡散という物理的な現象を表したものです。一方、「二酸化炭素と光合成」で説明したFarquharのモデル(簡単のために、低CO2での式のみ使います)は、CO2濃度(ここではCi)と光合成速度の生化学的な関係を表したものです。
二つの式は同じグラフの上で重ねることができます。P=gs(Ca-Ci) のグラフを裏返しにしてCO2-光合成速度関係のグラフにもってきます。ここで、P=gs(Ca-Ci) の原点(Ca-Ci=0、つまりCa=Ci)を、CO2-光合成関係のグラフ上のCa=Ciの点に重ねます(下の図参照)。
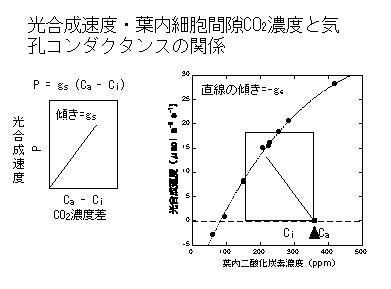
そうすると、P=gs(Ca-Ci) の式は、Ca=Ciの点から傾き-gsで上がっていく直線であることがわかります。この直線はどこかでCO2-光合成曲線と交わります。
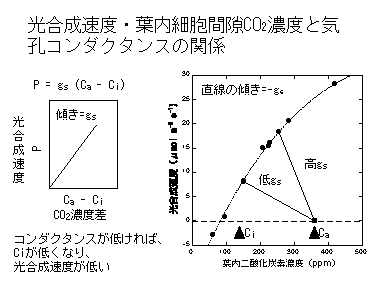
この交点こそが「実現される光合成速度」です。光合成の生化学的性質が一定でも、gsが高くなれば光合成速度が上がり、gsが低くなれば光合成速度が下がることがわかると思います。光合成速度はgsを上げるだけでも上がるし(下図C)、光合成系タンパク質(RuBPCase)量を増やしてVcmaxを上げることでも増えます(B)。もちろん、両方とも増やしたほうが効率よく増加することはいうまでもありません(D)。
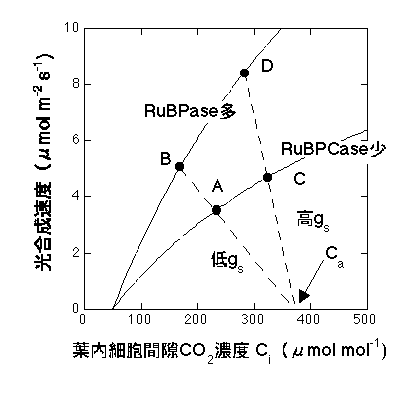
Farquhar and Sharkey (1981) はP=gs(Ca-Ci) を供給関数(supply function)、Farquharモデルで表されるCO2-光合成曲線を要求関数(demand function)と呼びました。つまり、光合成速度はCO2をどれだけ供給できるか、CO2をどれだけ必要とするか(CO2をどれだけの効率で光合成に使えるか、と考えてもよい)によって決まるということです。実現される光合成速度は二つの関数の連立方程式の解として得ることができます。
上に書いたように、光合成をするためにはCO2が必要で、気孔を開ければそれだけ光合成速度が上がることがわかります。しかし気孔は開けっ放しではなく、環境の変化に合わせてその開度が厳密に制御されています。なぜ気孔を閉じなくてはいけないか、というと、水のロスを防ぐためです。CO2が通りやすい、ということは、水蒸気が通りやすい、ということでもあります。CO2は欲しい、でも水は失いたくない、というジレンマから進化したのが気孔です。孔辺細胞はたいへん環境変化に敏感で、形態変化を伴うものとしては、最も環境応答が速いものの一つです(オジギソウがおじぎをするのには負けるかもしれません)。
気孔の開閉がどのようなメカニズムで行われているかは、様々な研究が行われ、かなりの部分が明らかになりつつあるようですが、私はよく知らないので省略します。
さて、気孔はどういうときに開いて、どういうときに閉じるのか、つまり気孔の開閉がどのように調節されているか、について書きましょう。簡単に書くと、以下のようになります。気孔コンダクタンスは・・・
・暗黒下ではほぼゼロ。光強度が高いほど高いが、ある程度以上の光強度では増加しなくなる。
・空気が乾燥すると下がる。
・CO2濃度が上がると下がる。
・土壌水分が不足すると下がる。
・葉の水ポテンシャルが下がると下がる。
・夜がくると(明るくても)下がる種もいる。
以上の応答にどのような意義があるかは別に詳しく述べますが、簡単にいうと、光合成しなくても良い条件では閉じる(暗黒下とか)、水を失うと困る条件では閉じる、といったところです。最後の「夜がくると」というのは、いわゆる体内時計による調節です。
空気の乾燥度が一定の場合、光やCO2濃度に対する変化は、おおざっぱには、「気孔の開閉はCi/Caが一定になるように調節されている」、といっていいのではないかというのが私の印象です。例えば、Caが変わると光合成速度も気孔コンダクタンスも大きく変化しますが、かなり低いCO2濃度を除き、Ci/Caの変化はあまりありません(Sage 1994, 下図)。
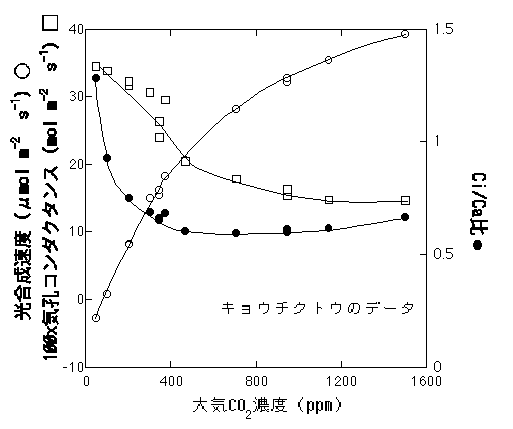
講義で図を作ったので、せっかくですから入れました。ちょっと大きいなあ。線はテキトー。
これを上の式を使って考えてみましょう。変形すると、
P/gs = Ca(1 - Ci/Ca)
となります。つまり、Caが同じなら、P/gsが同じになるように調節されているわけです。ですから、一般的に、種内でも種間でもPとgsの間には高い相関が見られます(報告多数、例えばWong et al. 1979, Yoshie 1986)。
大気の乾燥の程度が変わる場合は、「飽差」とCi/Caの関係を一定に保つように気孔の開閉が調節されているようです(Leuning 1995)。「飽差」とは、そのときの葉温における飽和水蒸気分圧(上で紹介した飽和水蒸気密度と同様の概念を密度ではなく分圧で表したもの)と実際の大気の水蒸気分圧の差で、Vapour pressure deficit(VPD)とよばれます。VPDが大きいほど空気が乾いていることを意味し、葉温と気温が同じならば、VPDはHi-Haと等しくなります。Leuningによれば、VPDとCi/Caの間には負の相関があり(VPDが大きいほどCi/Caが下がる)、この関係は気温の影響を受けません。ただし、VPDとCi/Caの関係は、一枚の葉においては保たれていますが、種間、生育条件の異なる葉の間では異なるようです。
さて、一日の中で気孔がどのように変化するかを考えてみましょう。まず、夜は気孔は閉じています。Ci/Caを一定にする、といっても、光合成速度が0や0以下になるような弱い光ではそもそもCiが下がらないので、この法則はあてはまりません。朝が来て、光が強くなってくると気孔が開いてきます。一般に、朝は気温が低いのでVPDが小さく、光が強くなるほど気孔が開いてきます(Pとgsが比例する)。昼になり、光が強くなると、気温が上がるためVPDが大きくなる可能性が出てきます。VPDがそれほど上がらない場合は、朝同様Pとgsは比例し、昼に光合成速度が最大になります。VPDが大きくなってしまうと、気孔は閉じます。乾燥地ですと、昼だというのに朝方よりもgsやPが低くなってしまうことは珍しくありません。このように、昼に気孔が閉じてしまうことを「昼寝現象」と呼びます(英語では単にmidday depressionと呼んでいるようです)。
Harley & Tenhunen (1991) はBall et al. (1987) が提唱したごく単純なモデルで日中の気孔閉鎖が説明つくことを示しています。Ballらのモデルは、ごく単純なものです。
gs = k P RH/Ca
RHは相対湿度(実際の水蒸気分圧を飽和水蒸気分圧で割ったもの)で、kは比例定数です(ただし、kは生育条件や種によって異なる)。この式を変形すると、
Ci/Ca = 1 - 1/k/RH
となります。Leuning (1995) ではVPDとCi/Caの間に負の相関が見られた、と書きましたが、これを式にすると、
Ci/Ca = a - b (VPD)
です。VPDと1/RHの関係は単調増加なので、この2つの式はおおざっぱには同じものです。Harleyの仕事はかなりきれいに気孔の挙動を説明しており、昼寝現象はVPDにせよ相対湿度にせよ、気孔と湿度の関係でほとんど説明できるのだろうと私は思います。
気孔の挙動のモデリングについては、古くはCowan (1977) からFarquhar & Wong (1984)、Ball et al. (1987) など多くの研究者によって試みが行われています。最近も検討は続いているようで、複数の論文を見た記憶があります。たぶんLeuning (1995) はその系列の論文の一本にすぎませんが、私は他の論文を読んだことがないので、ここではこの論文を挙げるにとどめました。この論文ばかり挙げるのは、この論文が示したデータ(VPDとCi/Caの関係は温度に影響されない、など)が非常にきれいで、強く印象に残っているからです。
以上は経験モデル、つまり気孔コンダクタンスが条件によってどのように変化するか、ということの積み重ねからどのように調節されているかを考えた研究です。これとは別に、どのように気孔コンダクタンスを調節すると有利か、という視点から最適な気孔コンダクタンスを予測した研究もあります。それは別に記すことにします。
「不均一な光合成」と言われてもなんのことだかわからないでしょうが、とにかくnon-uniform photosynthesis と呼ばれているのでこう訳すしかありません(Patchy photosynthesis と呼ばれることもあります)。何が不均一かというと、ある環境にある一つの葉の中で、気孔の開き方が不均一、つまり、開いている気孔と閉じている気孔が同時に存在しているということです。
まず、どうして気孔の開き方の不均一性が問題になるかを書きましょう。これは二酸化炭素と光合成の関係を考えるときに重要になります。上にはCiの計算方法を書きましたが、この計算方法は全ての気孔が同じように開閉することを前提としています。例を挙げて説明しましょう。
葉の表面に10個気孔がある植物を考えて下さい。葉の周囲の大気CO2濃度や光環境(飽和状態)は一定だとしましょう。乾燥か何かが起こって、徐々に気孔が閉じていくとします。全ての気孔が同様に閉じるときは、葉内細胞間隙のCO2濃度(Ci)は葉内のどの場所でも同じように下がり、そのため光合成速度も同じように下がります。このとき光合成速度とCiの関係はFarquharのモデルにしたがった減少をするはずです。気孔コンダクタンスが半分になれば、Ciや光合成速度も半分近く減少するはずです(半分とは限りません)。
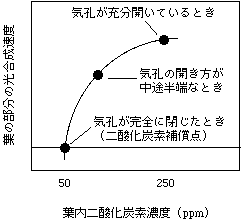
では気孔が不均一に閉じるケースを考えてみましょう。一番極端な例として、10個のうち5個の気孔は完全に開いていて、別の気孔が完全に閉じているケースを考えます。このとき、完全に閉じている気孔の近くの葉内細胞間隙では、光合成をしてもCO2が供給されないのでCO2濃度が大きく下がります。しまいには、CO2補償点まで下がってしまうでしょう。このときは光合成のCO2吸収と光呼吸のCO2放出が同量になっていて、見かけ上CO2吸収がありません。一方、気孔が開いている部分の葉内細胞間隙ではCO2供給があるので高いCiを維持することができます。気孔が完全に開いている場合と閉じている場合のCiをそれぞれ250、50ppmと仮定すると、葉全体のCiの平均は(5×250+5×50)/10で150ppmになります(下図白丸)。さて、光合成速度や蒸散速度を測ってみると、どうなっているでしょうか? 半分の気孔は元気に開いて、半分の気孔からは何も出てこない/入らないはずですから、光合成速度も蒸散速度はきっちり半分になります。蒸散速度が半分ですから、コンダクタンスも半分だと計算されます。このとき、Ciを計算してみましょう。P = gs(Ca - Ci) で、Caは一定、Pが半分、gsも半分ですが・・・Ciは変わりません。つまり、完全に閉じている気孔はP/gs比に影響を及ぼさないので、計算されるCiに影響がないのです。この計算を信用すると、Ciが変わらないのに光合成速度が落ちるという、摩訶不思議な状態になり、「Ci以外の原因で光合成速度が減少した!」という結論を導いてしまうわけです(下図黒丸)。
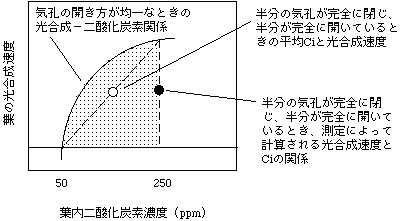
このような不均一な気孔コンダクタンスが起こる場合、光合成速度の二酸化炭素濃度依存性はFarquharモデルにしたがいません。Farquharモデルでは光合成速度とCiの関係は曲線ですが、平均Ciに対してプロットした例では光合成速度と葉の平均Ciの関係は直線になります。測定データを元に計算した場合は大きくFarquharモデルから逸脱し、モデルが正しいとして考えると、Vcmaxが減少したかのような錯覚に陥ります(Vcmaxについてはこちら)。もっと極端な例を考えると、10個の気孔のうち1個しか開かなかった場合は、Ciが変わらずに光合成速度が1/10になってしまいます。つまり、気孔の開き方によって図中網掛けの部分どこでも点が存在する可能性があるということです。
以上のような指摘を初めてしたのはどうやらLaisk (1983) らしいのですが、あいにく私は未読です。
さて、80年代当時、乾燥条件が引き起こす光合成速度の低下のメカニズムが話題になっていました。乾燥が気孔閉鎖を引き起こすことはほぼ間違いないのですが、葉内二酸化炭素濃度Ciと光合成速度の関係を見てみると、上で説明したように、単なるCiの低下だけでは光合成速度の低下を説明できないとする報告が多くありました。そこで、上で説明したような「不均一な光合成」で説明できるとする説(Laisk 1983)と、乾燥で光合成系がダメージを受けるのだ、という説がありました。
この問題を解決すべく行われたのがTerashima et al. (1988) の実験です。この実験では、まず1)気孔閉鎖時に、Ciの低下よりも光合成速度の低下が大きくなるのは実際に光合成系がダメージを受けたからなのか、を調べ、次に、2)実際に「不均一性」な気孔閉鎖が起こっているのかを調べました。
1)のほうは、従来通りのガス交換法(CO2と水蒸気の交換を同時に測定する)と、酸素電極法(こちら参照)の二つを併用することで調べました。酸素電極法では、葉を水を含んだスポンジの上に置きますので、けっこう葉に荒い処理をしても光合成速度を測定できます。寺島さんは表皮細胞を慎重にはがした葉の光合成速度を測定することで、光合成系がダメージを受けているのかを調べました。実験材料には、普通に気孔が開いている葉と、葉柄からアブシジン酸を吸わせた葉の比較を行いました。アブシジン酸(ABA)とは植物ホルモンの一種で、乾燥条件になると根で生産され、蒸散流にのって葉に達し、気孔を閉鎖する機能があることが知られている物質です。葉柄からABAを吸わせるということは乾燥条件をシミュレートし、気孔を強制的に閉じさせる処理なわけです。で、その結果ですが、ガス交換法で測定すると、ABAを吸わせた葉では上で説明したようなCiの低下よりも大きい光合成速度の低下が見られました。しかし、表皮をはがした葉の光合成速度を酸素電極で測定するとABAを吸わせた葉での光合成低下は見られませんでした。つまり、Ci低下以上の光合成速度の低下はあくまで見かけのものであり、実際には光合成系はダメージを受けていないことが示唆されました。
2)の「不均一性」のチェックはなかなか懐かしい方法を使って調べられました。それは(小学生でもできる)ヨウ素-デンプン反応です。寺島さんはABAを吸わせた葉をしばらく光にあて、その葉のデンプンの分布をヨウ素-デンプン反応で調べ、デンプン含量に大きく不均一性があることを示しました。これは葉の部分によって多く光合成できたところ(少し気孔が開いていた)とあまり光合成できなかったところがあることを意味します。
というわけで、Laisk (1983) が指摘したような不均一性が実際にあること、そしてその不均一性のせいで光合成速度-Ci関係が変わってしまうことが示唆されたわけです。以後気孔開度の不均一性は多くの研究によって存在が確認されています(後述)。ただし、後者の問題、つまり、乾燥時の光合成低下の原因が「不均一性」だけで説明できるのかについてはまだ結論がでていないようです。詳しくは、こちらに書くことにします。
homobaric leaf と heterobaric leaf
さて、また見慣れぬ言葉が出てきました。homobaric, heterobaricとも形態学用語です。ちょっと説明が前後してしまうのですが、不均一性の話をするときにこの話を除くわけにはいきませんので、少し寄り道して葉の形態の話をしましょう。
homobaric leaf , heterobaric leaf とも日本語訳がありまして、それぞれ「等圧葉」「異圧葉」といいます。使われてるのは見たことないですけどね。それぞれ何を意味するかというと、葉の内部形態が違います(下図)。
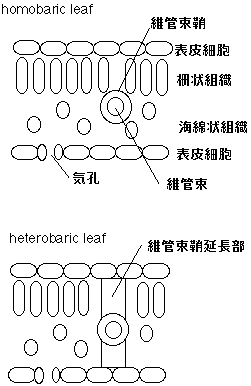
要するに、維管束鞘延長部があるか否かの違いです。heterobaric leafでは、この維管束鞘延長部があることにより、細胞間隙が分断されています。つまり、維管束鞘延長部に囲まれた部分は隔離された空間になっており、空気は維管束鞘延長部を越えて拡散することはできないのです。以後、この維管束鞘延長部で隔離された空間を「区画」と呼びます。
身近な植物ですと、ホウレンソウやソラマメなど葉脈があまり見えない葉はhomobaricであることが多いと思います。一方、ヒマワリやらオナモミなど葉脈が見える葉はたいていheterobaricだと思います。よくは知りませんが、たいていの種はheterobaric なのではないでしょうか。
どうしてこの話がここで出てくるかというと、もちろんheterobaric かhomobaricかによって、気孔開度の不均一性の影響が異なるからです。heterobaric の場合は、区画あたりの気孔は、少ないものでは10個程度だそうです。気孔開度の分布によっては、開いている気孔が一つもない区画もありえる一方、全てが開いている区画もあるかもしれません。そうなると、区画によって細胞間隙CO2濃度や光合成速度が大きく違うことになります。しかし、homobaric leafでは、ある気孔が閉じていても、遠くの開いている気孔を通してある程度のCO2供給ができ、不均一性はそれほど大きくないことが期待できます。上記のTerashima et al. (1988) ではhomobaric leaf のソラマメとheterobaric leaf のヒマワリを比較し、ABA処理されたヒマワリでは区画ごとにデンプン蓄積量が異なるのに対し、ソラマメでは不均一性が比較的小さいことが示されています。
このテーマは、KA Mottのグループを中心にかなり研究が進んでいます。いくつか論文を眺めてみて、だいたいこのテーマがどういう感じで進んでいるか書いてみたいと思います。私自身は水関係は専門外ですので、誤解があるかもしれないことをあらかじめ述べておきます。
○手法の進歩
Terashima et al. (1988) では、ヨウ素-デンプン反応というごく単純な方法を使って不均一性があることを示しました。同じ年にDownton et al. (1988) は14CO2のとりこみを使って不均一性があることを示しています。これらの方法の問題点は、測定において葉を破壊してしまうため、連続測定が不可能なことです。これでは気孔開度に不均一性があることはわかっても、その不均一性の時間変化はわかりません。そこで使われるようになったのが、クロロフィル蛍光を使った測定法です。クロロフィル蛍光を使った手法については、こちらに詳しく書きましたが、この研究で使われている手法はやや違って、クロロフィル蛍光強度の平面分布をモニタするものです(Daley et al. 1989)。この手法では、non-photochamical quenchingの二次元分布を測定します。non-photochemical quenching は光合成速度や気孔コンダクタンスと負の相関があるので、その相関を利用して区画の気孔コンダクタンスを推定しています。
この手法によってわかったことは、区画の気孔開度は必ずしも時間的空間的に一定ではないことです。例としてCardon et al. (1994) の研究を見てみましょう。彼らはヒマワリとオナモミ(どちらもheterobaric leaf)を使い、湿度低下後のクロロフィル蛍光の変動を数十分間観察しました(ヒマワリでは湿度低下とともに光強度も減少)。ヒマワリでは湿度低下とともに葉レベルの気孔コンダクタンスが「振動」しながら低下しました。つまり、単調に下がり続けるのではなく、波形のカーブを描きながら減少したのです。このときクロロフィル蛍光の平面分布を見てみると、最初からほとんど閉じっぱなしの気孔、開いたり閉じたりを繰り返す気孔が見られ、不均一性があることがわかりました。開いたり閉じたりの繰り返しは気孔間で比較的同調していました。一方、オナモミでは、葉レベルの気孔コンダクタンスは単調に減少し、各気孔の開度は変動していましたが、ヒマワリと違って同調していませんでした。
ちなみに、湿度が高いときには、光を弱くして気孔を閉じさせても不均一性は起こらないそうです。
○「不均一性」が起こるメカニズム
気孔を閉鎖させる実験手法として、Terashima et al. (1988) では人工的にABAを加えるという方法を、Downton et al. (1988) では鉢植え?の植物に水やりをやめ、水ストレスをかける、という方法を使っていました。後者の方法でも、気孔を閉鎖させるのはABAだと考えられています。寺島さんたちは当初気孔によってABAに対する反応が違うために不均一性が生じるのではないか、ということを考えていたようです(といいますか、少なくともTerashima et al. (1988) の実験ではそうだったはずです)。しかし、ABAが必ずしも不均一性を引き起こすために不可欠なわけではないことが明らかになりました。それは、ABAの生成を行わないトランスジェニック植物を用いた実験によって証明されています。Eckstein et al. (1998) はトランスジェニックタバコと野生型タバコの葉を植物体から切り離し、切り離し後の葉表面のクロロフィル蛍光分布を観察し、ABAがないトランスジェニックタバコでも野生型タバコでも不均一性を見いだしました。
現在のところ、不均一性が起こるメカニズムは、Mottらが精力的に調べています。Mottの興味は、単なる「不均一性」よりも、あるエリア内の気孔開度の変動が同調すること、つまり、周囲とは気孔開度が異なる「パッチ(patch)」がなぜ存在するのか、にあるようです。彼によれば、気孔開度の同調は表皮細胞と孔辺細胞の水力学的バランスによるものと考えられています。この説をうまく説明するのはなかなか難しいですが、次のようになります。気孔が開くためには、孔辺細胞の膨圧が上がることが必要ですが、孔辺細胞の膨圧が上がらなくても、孔辺細胞周辺の表皮細胞が縮めば、相対的に孔辺細胞が膨らむことになり、孔辺細胞の膨圧が上がらなくても気孔が開きます。湿度が急激に下がって蒸散速度が大きくなると、細胞から水が出ていくため、孔辺細胞・表皮細胞とも膨圧が下がります。たまたまある気孔が大きく開いて、周辺の表皮細胞の膨圧が大きく下がると、その膨圧が下がった表皮細胞の周辺にある孔辺細胞が大きく下がることになります。そうするとまた隣の孔辺細胞が・・・、というように、表皮細胞の膨圧の低下を通して周辺気孔の開き方が同調する、ということです。ただし、葉脈付近では、葉脈からの水供給があるため、葉脈を挟んで気孔開度が同調することはありません。この結果不均一性が起こるわけです。Mottらはこの仮説を証明するために様々な実験を行っているようです。私はフォローしていないのでよくわかりませんが(簡単なレビュー・・・Mott and Buckley 2000)。
○「不均一性」と光合成低下の関係
さて、乾燥にさらされた葉の光合成速度を低下の原因、特に、光合成-Ci関係が変化してしまうことの原因は何か?という問題にもう一度立ち返りましょう。Terashima et al. (1988) は、高CO2濃度での光合成速度を測定することにより、ABAを葉にかませても光合成系は傷まないことを示しました。彼らの結果の意味するところは、乾燥時の葉の光合成低下の原因は気孔開度の不均一性である、ということです。蛍光強度と光合成速度の関係から、Downton et al. (1988) も同様の結論を導いています。しかしこれに異を唱える論文が出されました。それはCheeseman (1991) の論文です。彼は実験をしたわけではなく、「本当に不均一性だけで光合成が低下するのか?」という問いに対し、コンピュータシミュレーションによって答えを探りました。彼はFarquharのモデル(こちら)を使い、様々な気孔閉鎖のパタンが起こったと仮定し、その葉全体の光合成への影響を調べました。その結果、気孔閉鎖の不均一性がベル型で表される場合は、不均一性が非常に大きいとしても光合成速度-Ci関係の変化はそれほど大きくはないはずだ、ということを理論的に示しました。つまり、上で挙げたような極端な不均一性(葉の気孔は全て閉じているか、全て開いているかのどちらか)が起こっているのでなければ、不均一性だけでは光合成速度はそれほど下がらない、ということです。
上に書きましたように、90年代に入ってから、蛍光を使って不均一性を調べる研究が増えています。これらの結果を見る限りでは、気孔は全て閉じているか、全て開いているかのどちらか、ということはないようです(ただし、使われている材料には限りがありますが)。したがって、Cheesmanがいうように、不均一性だけで光合成低下を説明できないことになります。しかし、ではTerashima et al. (1988) が示した結果はどう考えるんだ、というと、Cheeseman はそれに対する答えは何も考察していないので、わからないとしか言いようがありません。
Mott (1995) は「不均一」の程度の大きさと光合成-Ci関係の変化の間には相関があると言っているようですが、あいにく未読です。
Eckstein et al. (1998) の論文では、乾燥にさらした葉の光合成速度はCi一定のまま低下します(見事に一直線です)。彼らはこの低下の原因は「不均一性」と葉肉細胞でのなんらかのダメージの両方であろう、という言い方をしています。彼らの言い方からすると、98年の時点ではわかっていないようですね。
Terashima et al. (1988) の実験結果に対しては、一つ問題点を指摘することができます。彼らは表皮細胞をはがして、高CO2濃度での光合成速度を測定し、光合成系が傷んでいないことを示しました。しかし、こちらで説明しましたように、高CO2ではRuBPCaseの活性が低下していたとしら、それは感知できません。したがって、彼らの結果では、RuBPCase活性が低下していた可能性は否定できないのです。ただ、乾燥にさらした葉でRuBPCaseが不活性化する、という話はあまり聞いたことがありません。私が持っている論文の中では、Vu and Yelenosky (1988) が、光合成速度が日中低下するとき(昼寝現象)でもRuBPCaseの活性化レベルは下がらないことを示しています。
別の可能性として、葉内細胞間隙から葉緑体へのCO2拡散が悪くなるのではないか、ということが考えられます。このCO2拡散については、詳しくは下で書きます。乾燥条件とはちょっと違う話である可能性がありますが、Delfine et al. (1998) は塩ストレスにさらされた植物の葉では葉内細胞間隙から葉緑体へのCO2拡散が悪くなり、葉緑体内のCO2濃度が下がるせいで光合成速度が下がることを示しています。ひょっとしたら、乾燥時もこういったことが起こっているのかもしれません。
最後に一言。この問題に私に多少なりとも知識があるのは寺島さん(大阪大学)のおかげです。Delfineの論文も寺島さんに紹介してもらいました。