熊野研究室 発生生物学分野
東北大学大学院生命科学研究科付属浅虫海洋生物学教育研究センター
| トップページ | 研究内容 | メンバー | 業績 | 論文セミナー | 連絡先 | アクセス | リンク | 浅虫海洋生物学教育研究センター |
トップページ > 研究内容
<はじめに>
2013年7月にできた研究室です。
浅虫周辺に生息する海産無脊椎動物を用いて、以下のような発生生物学的研究を行っています。
1)海洋プランクトンの体の形づくりの仕組みの解析
2)生殖細胞系列形成機構の解析
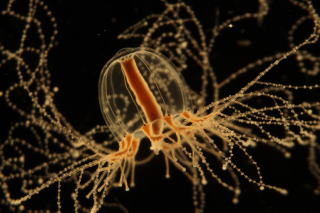 地球上には既知の動物だけでも140万種ほど存在し、それぞれが固有の形をした体や器官を持ちます。このような多様な形が、進化の過程でどのように生じてきたのかを理解することは、生物学における最重要課題の一つです。多様な形が存在するのは、形をつくる仕組みが動物種ごとに異なるからだと考えられます。近年の研究により、動物における体や器官の形づくりの仕組みについて多くの知見が得られていますが、そのほとんどは、哺乳類を中心とした脊椎動物やショウジョウバエを対象にした研究成果に基づきます。我々は、これまであまり研究のされていない動物種を用いて、動物の形づくりにおける新しい仕組みを明らかにしようとしています。現在、注目しているのは、刺胞動物エダアシクラゲ(Cladonema pacificum)の触手が枝分かれする過程と、脊索動物マボヤ(Halocynthia roretzi)のオタマジャクシ幼生が、胴部と尾部の境界を「くびれ」としてはじめて形成する過程です。
地球上には既知の動物だけでも140万種ほど存在し、それぞれが固有の形をした体や器官を持ちます。このような多様な形が、進化の過程でどのように生じてきたのかを理解することは、生物学における最重要課題の一つです。多様な形が存在するのは、形をつくる仕組みが動物種ごとに異なるからだと考えられます。近年の研究により、動物における体や器官の形づくりの仕組みについて多くの知見が得られていますが、そのほとんどは、哺乳類を中心とした脊椎動物やショウジョウバエを対象にした研究成果に基づきます。我々は、これまであまり研究のされていない動物種を用いて、動物の形づくりにおける新しい仕組みを明らかにしようとしています。現在、注目しているのは、刺胞動物エダアシクラゲ(Cladonema pacificum)の触手が枝分かれする過程と、脊索動物マボヤ(Halocynthia roretzi)のオタマジャクシ幼生が、胴部と尾部の境界を「くびれ」としてはじめて形成する過程です。

次世代をつくる卵と精子は、生殖細胞系列の細胞(始原生殖細胞)としてその運命が決定し、その他の体細胞系列の細胞とは運命を別にして発生します。このように、全能性をもち、唯一次世代に受け継がれる特殊な細胞をつくる生殖細胞系列を発生中に確立・維持することは、 種を存続するうえで生物にとって必須のイベントであり、それゆえその仕組みは進化の過程で容易には変化してこなかったのだと考えられてきました。ところが近年の幅広い動物種を用いた研究により、実は生殖細胞系列を形成する仕組みは多様であることがわかってきました。我々は、脊索動物ワカレオタマボヤ(Oikopleura dioica)の生殖細胞系列形成機構を解析することで、同じ尾索動物であるホヤのそれとの比較により、この多様な仕組みがどのように生じてきたのかを明らかにしようとしています。また、これまでこの分野であまり研究がなされてこなかった刺胞動物における生殖細胞系列形成の仕組みを解析することで、より幅広い動物種での本仕組みの多様性(または共通性)について明らかにしようとしています。さらに、ホヤは始原生殖細胞が再生することが知られる珍しい動物です(他にウニと多毛類Capitellaで知られるのみ)。したがってこの再生を制御する遺伝子制御ネットワークを明らかにしようとしています。
種を存続するうえで生物にとって必須のイベントであり、それゆえその仕組みは進化の過程で容易には変化してこなかったのだと考えられてきました。ところが近年の幅広い動物種を用いた研究により、実は生殖細胞系列を形成する仕組みは多様であることがわかってきました。我々は、脊索動物ワカレオタマボヤ(Oikopleura dioica)の生殖細胞系列形成機構を解析することで、同じ尾索動物であるホヤのそれとの比較により、この多様な仕組みがどのように生じてきたのかを明らかにしようとしています。また、これまでこの分野であまり研究がなされてこなかった刺胞動物における生殖細胞系列形成の仕組みを解析することで、より幅広い動物種での本仕組みの多様性(または共通性)について明らかにしようとしています。さらに、ホヤは始原生殖細胞が再生することが知られる珍しい動物です(他にウニと多毛類Capitellaで知られるのみ)。したがってこの再生を制御する遺伝子制御ネットワークを明らかにしようとしています。
<研究内容>
1)エダアシクラゲ触手枝分かれ機構の解析
動物の組織や器官はその機能に適した固有の形を採り、その形成過程では、最外層の上皮細胞と裏打ちする中胚葉細胞との相互作用が重要であることが知られています。特徴的な器官形態の1つである枝分かれについても、近年の哺乳類やショウジョウバエの研究から、中胚葉からのReceptor
Tyrosine Kinase (RTK)シグナルによる上皮シートの変形によることわかってきています。我々はこれまでに、中胚葉を持たないに二胚葉性動物であるエダアシクラゲの枝分かれした触手を研究材料にして、上皮・中胚葉相互作用に依存しない動物における新たな形づくりの仕組みを明らかにしてきました(Fujiki et al., 2019; Hou et al., 2021)。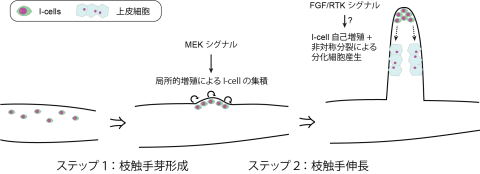 すなわち、触手枝分かれは、クラゲ傘から伸びる主触手が成長するに伴い、その基部で次々と新しい枝触手ができること、ヒドロ虫綱に特有の多能性幹細胞であるinterstitial
cell (I-cell)が、枝触手形成に先立って主触手上の枝分かれ部位に枝形成ごとに繰り返し集積してくること、集積したI-cellからは枝触手を構成する上皮細胞など複数の細胞タイプが生じてくること、この枝触手形成・伸長過程にRTKシグナルが必要であること、などを明らかにしました。これらの結果により、二胚葉性動物の形づくりにおいては、多能性幹細胞が中心的な役割を果たすことがわかりましたが、同時に、一見複雑な枝分かれ構造も単純なルールの繰り返しによりつくられることや、RTKシグナルが必要であることなど、広く動物界にわたって共通の仕組みが利用されていることも確認できました。現在は、1)繰り返し起こる触手枝分かれがどのように時空間的に制御されているのか、2)RTKシグナルにより多能性幹細胞の機能がどのように制御され枝触手形成に寄与しているのか、3)触手の枝分かれという新奇形質が進化の過程でどのように獲得されたのか、を理解すべく解析を進めています。
すなわち、触手枝分かれは、クラゲ傘から伸びる主触手が成長するに伴い、その基部で次々と新しい枝触手ができること、ヒドロ虫綱に特有の多能性幹細胞であるinterstitial
cell (I-cell)が、枝触手形成に先立って主触手上の枝分かれ部位に枝形成ごとに繰り返し集積してくること、集積したI-cellからは枝触手を構成する上皮細胞など複数の細胞タイプが生じてくること、この枝触手形成・伸長過程にRTKシグナルが必要であること、などを明らかにしました。これらの結果により、二胚葉性動物の形づくりにおいては、多能性幹細胞が中心的な役割を果たすことがわかりましたが、同時に、一見複雑な枝分かれ構造も単純なルールの繰り返しによりつくられることや、RTKシグナルが必要であることなど、広く動物界にわたって共通の仕組みが利用されていることも確認できました。現在は、1)繰り返し起こる触手枝分かれがどのように時空間的に制御されているのか、2)RTKシグナルにより多能性幹細胞の機能がどのように制御され枝触手形成に寄与しているのか、3)触手の枝分かれという新奇形質が進化の過程でどのように獲得されたのか、を理解すべく解析を進めています。
2)マボヤ神経胚「くびれ」形成機構の解析
動物の体や器官の形づくりにおいてよく観察され、形づくりの基本的メカニズムとして知られる「上皮の屈曲」(epithelial bending)は、様々な動物における研究により、頂端収縮や分裂に伴う細胞の円形化などが要因となっていることが明らかになっています。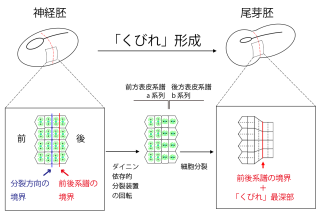 我々はこれまでに、マボヤ神経胚側方部の将来の胴尾部境界に「くびれ」が出来る過程で、上皮屈曲の新たな仕組みを発見しました(Nakamoto and Kumano, 2020)。すなわち、神経胚最外上皮層を構成する表皮細胞において、前方と後方で明確な境界を示しつつ互いに垂直となる異なる方向で(前方:胚周囲に沿った方向、後方:胚前後に沿った方向)細胞分裂をし、結果的に分裂後の胚周囲に沿った細胞数または周囲長が前後で異なることで「くびれ」が形成される、というものです。この分裂方向の違いは、後方の表皮細胞において、分裂直前に分裂装置が回転し胚の前後軸に沿った方向に配向することでもたらされます。現在は、1)後方表皮細胞において分裂装置の回転がどのような分子の作用で起こるのか、2)胚の後方だけでなぜ分裂装置の回転が起こるのか、3)分裂方向の境界の胚の前後軸に沿った位置がどのように決められているのか、を理解すべく解析を進めています。
我々はこれまでに、マボヤ神経胚側方部の将来の胴尾部境界に「くびれ」が出来る過程で、上皮屈曲の新たな仕組みを発見しました(Nakamoto and Kumano, 2020)。すなわち、神経胚最外上皮層を構成する表皮細胞において、前方と後方で明確な境界を示しつつ互いに垂直となる異なる方向で(前方:胚周囲に沿った方向、後方:胚前後に沿った方向)細胞分裂をし、結果的に分裂後の胚周囲に沿った細胞数または周囲長が前後で異なることで「くびれ」が形成される、というものです。この分裂方向の違いは、後方の表皮細胞において、分裂直前に分裂装置が回転し胚の前後軸に沿った方向に配向することでもたらされます。現在は、1)後方表皮細胞において分裂装置の回転がどのような分子の作用で起こるのか、2)胚の後方だけでなぜ分裂装置の回転が起こるのか、3)分裂方向の境界の胚の前後軸に沿った位置がどのように決められているのか、を理解すべく解析を進めています。
3)海産無脊椎動物の生殖細胞系列形成機構の解析
動物の生殖細胞系列形成には大きく分けて4つの方法、すなわち、Preformation型(例、ショウジョウバエ、線虫、カエル、ホヤ)、Epigenesis型(例、マウス、サンショウウオ、コオロギ)、After
embryogenesis型(例、貝、環形動物)、Adulthood型(例、刺胞動物、プラナリア)があり、その機構は多様であることが知られています。我々はこれまでに、マボヤから生殖細胞系列におけるグローバルな転写抑制を制御する母性局在因子PEM(Posterior
End Mark)を同定し、Preformation型を採る動物の間でも、生殖細胞系列の維持機構に分子レベルで多様性が見られることを明らかにしてきました(Kumano et al., 2011)。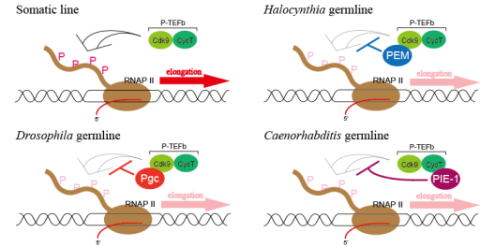 PEMは、ハエと線虫で同定されたそれぞれの母性因子PgcとPIE-1と同様に、pTEFbという複合体に結合しRNA polymerase IIのリン酸化を抑制することでグローバルな転写抑制を行いますが、これら3因子はそれぞれの系統にしか存在しない進化的に新しい因子で、それぞれにユニークな母性局在因子が、pTEFbを介したRNA
polymerase IIのリン酸化抑制という共通の機構で生殖細胞系列での転写を制御すると示唆されています。ホヤと、ハエ・線虫は系統的に離れており、それぞれ独立にPreformation型を獲得したと考えられていますが、ホヤと共通のPreformation型の祖先をもつオタマボヤにおいても、そのゲノムにはPEMが存在しないことがわかっています。現在は、1)進化の過程でこのような多様な制御機構がどのように生じてきたのかを理解するために、オタマボヤの生殖細胞系列において転写を抑制する因子の同定および転写制御機構の解析を進めています。
PEMは、ハエと線虫で同定されたそれぞれの母性因子PgcとPIE-1と同様に、pTEFbという複合体に結合しRNA polymerase IIのリン酸化を抑制することでグローバルな転写抑制を行いますが、これら3因子はそれぞれの系統にしか存在しない進化的に新しい因子で、それぞれにユニークな母性局在因子が、pTEFbを介したRNA
polymerase IIのリン酸化抑制という共通の機構で生殖細胞系列での転写を制御すると示唆されています。ホヤと、ハエ・線虫は系統的に離れており、それぞれ独立にPreformation型を獲得したと考えられていますが、ホヤと共通のPreformation型の祖先をもつオタマボヤにおいても、そのゲノムにはPEMが存在しないことがわかっています。現在は、1)進化の過程でこのような多様な制御機構がどのように生じてきたのかを理解するために、オタマボヤの生殖細胞系列において転写を抑制する因子の同定および転写制御機構の解析を進めています。
またAdulthood型を採る動物では、生殖細胞を含むほぼ全ての細胞タイプに分化する能力を持つ多能性幹細胞が成体まで維持され、成体期に多能性細胞から恒常的に生殖細胞や体細胞が作り出されています。この際どのようにして生殖細胞と体細胞の別を生み出しているのかはほとんどわかっていません。そのため我々は、現在、エダアシクラゲを用いて多能性幹細胞I-cellから生殖細胞へ分化する細胞が生じる過程に注目し、その制御機構を明らかにしようとしています。
最後に、ホヤのオタマジャクシ幼生では尾の先端部に始原生殖細胞が存在しています。この幼生の尾の先端部を切除して始原生殖細胞を除去した幼生は、そのまま変態をし成体へと発生しますが、驚くべきことに受精可能な生殖細胞ができてくることが知られています。現在、1)ホヤ幼生がどのようにして始原生殖細胞を失ったことを感知し再生しはじめるのか、2)その制御はどのような分子の作用で起こるのか、3)どの細胞タイプがどのようにして始原再生殖細胞へと転換するのか、を理解すべく解析を進めています。