光合成の生理生態学講座
目次
はじめに(200007 06)
光合成誘導(200007 13)
弱光から強光へ(200007 21)
強光から弱光へ(200007 27)
光利用効率(200008 17)
「光と光合成」で書いてきた光と光合成の関係については、多くの場合「定常状態」を仮定しています。ここでいう「定常状態」というのは、同じ強さの光を当て続けており、光合成速度が安定している状態のことを言います。しかし野外では太陽の移動により光強度は刻々と変化しています(こちら)。特に、雲や上層の葉による遮蔽は光強度の大きな変化をもたらします。定常状態では、光強度と光合成速度の間には一定の関係がありますが、「非定常状態」では、この関係が崩れます。この項では、非定常状態での光合成速度について書きます。
光強度が突然増加したときの光合成速度の応答は、その前に葉がどのような環境にいたかによって大きく異なります。一般に、暗黒下や弱光下では、酵素が不活性化していたり、気孔が閉じ気味になっていたりで、光が突然当たってもすぐに光合成できるわけではありません。いわば、光合成系は「眠って」いるわけです。眠っている葉に強い光を当てると、酵素の活性化などが徐々に起こり、光合成速度が徐々に増加します。光強度を上げてこの眠りから「目覚める」ことを「誘導」(induction)と呼びます。
1 光活性化酵素
光合成系酵素では、光が当たっている状態でないと活性化される酵素がいくつかあります。ATPase、NADP-G3PDase、FBPase、SBPase、Ru5P kinaseなどです。これらの酵素の活性化のメカニズムが全て同じなのかは私はよく知りませんが、少なくとも、FBPaseは以下のようにして活性化されます。光があたり、電子伝達反応が起こります。電子はフェレドキシンからNADPに伝達されるわけですが、一部の電子はフェレドキシンからチオレドキシンという物質に渡されます。さらに、チオレドキシンからFBPaseに電子が渡ります(もう少し介在電子伝達物質があったかも)。不活性型FBPaseはジスルフィド結合(-S=S-)を持っていますが、電子を受け取ることによりこの結合が切れ、活性型になります。
2 RuBPCase
RuBPCaseも不活性化される酵素です。この酵素の活性化・不活性化にはいくつか方法があり、種によって異なります。
i)CO2とMg2+
CO2とMg2+がリジン残基に結合することによりRuBPCaseが活性化されます。
ii)CA1P(carboxyarabinitol-1-phosphate)
CA1Pはリン酸を一つもつ糖です。暗黒下で生成され、RuBPCaseの活性部位に結合し、不活性化させます。CA1Pは全ての植物にあるわけではなく、インゲンはCA1Pで活性調節されているようですが、ホウレンソウにはCA1Pがありません。
iii)他、RuBPなどの物質が活性部位に結合して不活性化することもあるようです。
これらの原因により不活性化されたRuBPCaseはrubisco activaseと呼ばれる酵素によって活性化されます。酵素の活性化のしくみについてはいずれどこかで詳しく書きたいと思います。
3 気孔コンダクタンス
気孔は一般に弱光では閉じ気味で、光が強くなると開きます。気孔が開くとともに気孔コンダクタンスが増加します。
誘導されていない葉に強い光をあてると、1-2分の間に光合成速度は急速に上昇します(fast-phase)。その後は時間と共にゆっくり光合成速度が上昇します(slow-phase)。Fast-phaseではFBPaseなどの光活性化酵素の活性化が光合成速度を律速していると考えられています(Kirschbaum and Pearcy 1988b, Sassenrath-Cole and Pearcy 1992, 1994)。Slow-phaseでは、RuBPCaseの活性化だと言われています(Pearcy 1988, 1990)。気孔コンダクタンスですが、乾燥している場合は気孔コンダクタンスの影響が大きいようですが、湿潤な場合は小さいようです(Tinoco-Ojanguren and Pearcy 1993)。林床は湿潤な場合が多いので、あまり問題にはならないと考えられています。
では、すでに誘導してある葉では、弱光から強光に移したときに、どういうことが起こるでしょうか。
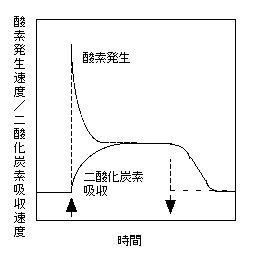
下の図は、数十umol m-2 s-1の弱い光があたっている葉に数百umol m-2 s-1の強い光を10秒ほどあてたときの、酸素発生速度と二酸化炭素吸収速度の模式図です。酸素発生は光が強くなるとともにすぐ増大します。ピーク時には定常状態での速度をはるかに上回りますが、すぐに下がります。また、光がまた弱くなると、酸素発生速度は直ちに低くなります。一方、二酸化炭素吸収速度は、徐々に(といっても実際は短い時間ですが)速度が上がり、定常状態に達します。光を消してもすぐには吸収速度は下がりません。
この図はKirschbaum and Pearcy (1988a) の図を元にしています。この業界ではなかなか有名な図です。
瞬間的には酸素発生速度と二酸化炭素吸収速度は一致していません(定常状態では一致します)。しかし、積算で見ると、強い光があたったことによる酸素発生量と二酸化炭素量はおおよそ同じです。
この「decoupling」は、光合成反応がどういうメカニズムかを考えると理解できます。光化学系IIでは光があたると、ほんの短い時間で水の分解・酸素の発生が起こります。しかし、二酸化炭素吸収は、光があたって、電子が伝達されて、ATPと還元力が生産されて、カルビンサイクルが回って、はじめて二酸化炭素が吸収されます(なお、誘導されている葉を暗い場所に置いておくとPGAが蓄積されています)。光が弱くなったあとも同様で、酸素発生は光が弱くなると直ちに速度が低くなりますが、二酸化炭素吸収のための中間代謝産物はまだ葉の中にあるので、消光後もある程度の同化を行うことができるわけです。
なお、光を強くした直後の高い酸素発生速度は、光化学系の電子伝達能力が、その下流の電子伝達構成要素の能力より高いために起こります。具体的に言うと、弱光下ではプラストキノンの多くは酸化型になっています。急に強光になると、光化学系IIは電子をプラストキノンに伝達します。このときは酸化型が多いので高い速度になります。しかし、ある程度電子が伝達され、還元型のプラストキノンが増えると、伝達する相手が減るので、電子伝達速度が制限されてしまうわけです。
次は、強光から弱光に移ったときです。
光を弱くした場合、強光をどれだけ長い時間を当てていたかでCO2吸収速度の変化が異なります。下の図は、数十umol m-2 s-1の弱い光があたっている葉に数十umol m-2 s-1の強い光をあて(上向き矢印)、再び弱い光に戻したとき(下向き矢印)の、二酸化炭素吸収速度の模式図です。20秒程度しか当てていなかった場合(下図点線)、上記のように、光を弱くしたのち、光合成速度は徐々に下がります。光を弱めてもなおCO2を吸収する現象を、Post-illumination CO2 assimilationといいます。これは、上で説明したように、蓄積した中間代謝産物をゆっくり利用しているわけです。
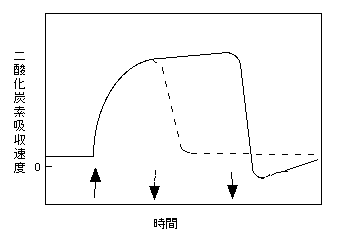
Peacy (1990) より。
一方、強光を1分以上あてていると、CO2吸収速度の減少は負になる、つまり、CO2の放出が起こります(上図実線)。これをpost-illumination CO2 burstと呼びます。なお、CO2放出は長く続くわけではなく、一時的なものです。このCO2放出には、光呼吸が関わっていることが知られています。例えば、酸素濃度を2-3%まで減らしてやると、消光直後のCO2放出が大きく減ります(Azcon-Bieto 1983)。また、光呼吸が起こらないC4植物でも同様です(Byrd et al. 1992)。これは、カルビンサイクルの一回りよりも、光呼吸のグリコール酸回路の一回りのほうが時間がかかるため、中間代謝産物の消費が遅れて現れることによります。ちなみに、この現象が光呼吸の発見につながったようですね。
どれだけ効率よくサンフレックを利用しているか、ということを定量化するため、光利用効率light use efficiency(以下LUE)というパラメータが提案されています(Chazdon and Pearcy 1986)。仮に、光強度の変化にタイムラグなしに対応し、瞬間的に定常状態に達することができる葉を考えます。この仮想的な葉がサンフレックによって吸収する二酸化炭素量(図中イ)に対し、実際の葉がサンフレックによって吸収した二酸化炭素量(図中ア)を比較します。このア/イ比をLUEと定義するわけです。
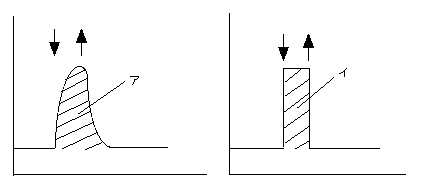
縦軸は光合成速度、横軸は時間。弱光を当てている葉に突然強光を短時間当てたとき(矢印)の応答
LUEはサンフレックの持続時間や光合成系がどれだけ誘導されているかによって違います。インドクワズイモの結果(Chazdon and Pearcy 1986)をご紹介しましょう。充分誘導されている場合、5秒間のサンフレックでのLUEは約1.6、つまり、定常状態よりはるかに効率がよい二酸化炭素吸収をします。これは、上に少し書いたように、光化学系IIの電子伝達能力が高いことによると思われます。つまり、短いサンフレックの間なら、下流の消費能力を上回る光化学反応を行い、プラストキノンプールを一気に還元し、光が消えた後もその還元力をゆっくり利用できるためです。しかし、サンフレックの時間が長くなると、この効果はなくなります。40秒のサンフレックでは、LUEはほぼ1になります。
これに対し、全く誘導されていない場合は、全体的にLUEが低くなります。5-10秒のサンフレックでようやくLUEが1、40秒のサンフレックでは0.6程度に落ちてしまいます。
ChazdonとPearcyは、同じ誘導レベルで比べると陽生植物のLUEは陰生植物のLUEより低い、といっています。これは、陰生植物がサンフレックをより有効に利用している、ということを示しているのかもしれません。種間差については、Tinoco-Ojanguren and Pearcy (1992) などが調べています。今のところ、陰生植物のほうが気孔の開閉がサンフレックに対して敏感であるなどの知見が得られているようです。