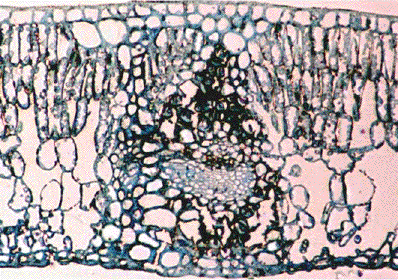
ヤブツバキの断面写真。一度だけ自分でとった断面写真のうち、唯一なんとか人に見せられるレベルにあるものです。
光合成の生理生態学講座
※改訂途中のため一部リンクが切れています。
目次
はじめに(200402 12)
葉の内部構造(200402 12)
葉内の光の分布(200402 25・200405 22)
葉緑体運動(200403 15)
葉緑体の順化(200404 07)
葉内のCO2拡散(200405 07・200406 04)
高い光合成速度を持つために葉がしなければいけないこと(200406 04)
光合成をする単位、というのを考えると、最小の単位は葉緑体で、最大の単位は森林などの植物群集ということになるでしょうか。我々生理生態レベルの研究者は葉(あるいは葉面積)を光合成の単位として扱います。光合成速度を葉面積あたりで表すときには、暗に葉を光合成する固まりとして考えているわけですが、葉の光合成は葉の中に含まれる葉緑体の光合成の和です。葉の中の葉緑体はその位置にしたがって微環境も性質も異なります。ここでは、葉が光合成をしているときに、葉の中では何が起こっているのか、葉緑体の視点から見てみることにしましょう。
まずは基礎知識として葉の内部構造を見てみることにしましょう。
ヤブツバキの断面写真。一度だけ自分でとった断面写真のうち、唯一なんとか人に見せられるレベルにあるものです。
双子葉植物に普通に見られる背腹葉(dorsiventral leaf、上面から光を受けるような平たい葉)を例に説明します。葉の上面(形態学用語では向軸側adaxialという)と下面(背軸側abaxial)は表皮細胞(epidermis cell)で覆われています。多くの種では表皮細胞は1層ですが、多層の表皮を持つ種もいるようです。表皮細胞の外側はクチクラ(ワックス)で覆われていて、水分などが蒸発しないようになっています。通常表皮細胞には葉緑体はありません。例外は、気孔を形成する孔辺細胞(guard cell)です。
多くの植物は下面のみに気孔を持ちます(下面気孔性、hypostomatic)が、両面に気孔を持つ植物(両面気孔性、amfistomatic)も少なくありません。まれに、水生植物など上面にのみ気孔を持つ植物もあります(上面気孔性、epistomatic)。
表皮細胞の内側には葉肉細胞があります。葉肉細胞内には多数の葉緑体があり、光合成を行っています。表皮細胞の内側は細胞がつまっているわけではなく、細胞と空間が入り交じっています。この空間は細胞間隙(あるいは空隙、intercellular space)と呼ばれ、液体ではなく気体で満たされています。
多くの葉では葉肉細胞には形態的な分化が見られます。表側に位置する葉肉細胞は細長い形をしており、柵状組織細胞(palisade cell)と呼ばれます。裏側に位置する葉肉細胞は海綿状組織細胞(あるいは柔組織細胞, spongy cell)と呼ばれます。海綿状組織細胞は、上の写真のような断面図では丸く見えますが、実際には木の枝のようにあちこちに手を伸ばす複雑な形をしています(Lambers et al. 1998 Plant Physiological Ecologyをお持ちの方は、11ページをご覧下さい)。
表皮の内側にあるのは葉肉細胞だけではなく、維管束(vascular bundle)もあります。維管束は必ず維管束鞘(bundle sheath)で覆われています。維管束鞘が発達し、細胞間隙を分断している葉を異圧葉(heterobaric leaf)、維管束鞘が貧弱で、維管束の左右の細胞間隙が連絡している葉を等圧葉(homobaric leaf)といいます。
ここまで背腹葉を例に書きましたが、他に両面葉(bifacial leaf)、双同側形葉(isobilateral leaf)、単面葉(unifacial leaf)とかいうのがあるらしいですが、どう違うのかよくわかりません。
以上、ルダル(1997)植物解剖学入門を参考にしました。
付録:葉にまつわる形態学用語とその意味
|
abaxial |
背軸側 |
茎の反対側を向いている葉表面 |
|
adaxial |
向軸側 |
茎のほうを向いている葉表面 |
|
amfistomatic |
両面気孔性 |
|
|
bifacial leaf |
両面葉 |
|
|
bundle sheath |
維管束鞘 |
|
|
dorsiventral leaf |
背腹葉 |
|
|
epidermis cell |
表皮細胞 |
|
|
guard cell |
孔辺細胞 |
|
|
heterobaric leaf |
異圧葉 |
細胞間隙が分断 |
|
homobaric leaf |
等圧葉 |
細胞間隙が連絡 |
|
hypostomatic |
下面気孔性 |
|
|
intercellular space |
細胞間隙 |
|
|
palisade cell |
柵状組織細胞 |
|
|
spongy cell |
海綿状組織細胞 |
|
|
vascular bundle |
維管束 |
|
葉は私たちの目から見ると非常に薄いものですが、光学的にはその厚さは無視できません。葉の内部にも光環境の勾配があり、背腹葉ならば向軸側(表)に近い側の葉緑体ほど強い光を受けることになります。一般に、葉の透過率(向軸側から当てた光のうち背軸側から出てきてしまう光の割合)は5-10%程度ですから、最も向軸側の葉肉細胞と最も背軸側の葉肉細胞では10倍程度の光環境の違いがあることになります。
葉内の光を考える上で、まず重要なのは、葉緑体の存在です。葉緑体は多数のクロロフィル分子を持ち、当たった光をかなりの確率で吸収します。向軸側の葉緑体が入ってきた光を優先的に吸収すれば、必然的に背軸側へ到達する光の量は少なくなります(群落内で下部ほど暗くなるのと同じ理屈です)。また、クロロフィルは青と赤に吸収極大があり、緑の光はあまり吸収しません。このため波長によって光強度の分布が違う(緑の光は背軸側まで達しやすい)ことになります。
次に重要なのが細胞間隙の存在です。細胞間隙は上述の通り気相で、細胞は実質上液相であるとみなすことができます。前者の光の屈折率は1.0(屈折しない)で、後者の屈折率は1.5です(寺島 2002)。このため、光が細胞と細胞間隙の境界を通過するたびに光の屈折が起こることになります。屈折とはつまり光の方向を変えることで、方向が変わることにより、光は葉内を散乱することになります。このような散乱は、「光が葉緑体に吸収される確率」を高めます。屈折がなければそのまま葉を透過していた光が散乱によって葉に留まるわけです。葉内に留まる確率が高ければ、それだけ吸収されるチャンスも多いわけです。
この散乱の重要性は簡単に実験で示すことができます。注射器の中に葉の切片と水を入れ、注射器の入り口をふさいでピストンを何度も往復させると、注射器内の圧力の急激な変化をきっかけとして細胞間隙に水が入り込みます(infiltrationと呼びます)。普通の葉と細胞間隙内が完全に水で満たされた葉の光の透過率を比較すると、後者のほうが10%程度透過率が高くなります(Moss & Loomis 1952)。細胞間隙内が水で満たされ、屈折が起こらなくなったからです。
柵状組織と海綿状組織の細胞の形の違いにも重要な意味があります。柵状組織は円筒形をしていて光を通しやすい形になっていますが、海綿状組織は複雑な形をしているので屈折が起こりやすくなり、光を吸収するチャンスが多くなります。柵状組織はもともと強い光が当たるので、わざわざ吸収効率を高める必要がありません。一方海綿状組織では柵状組織を通過した弱い光しかあたりません。つまり形態の違いは、向軸・背軸側の光環境の不均一性を是正するような適応と考えることができます(Terashima and Saeki 1983)。
細胞内の葉緑体の配置も葉内の光分布に影響を与えます。後で述べるように葉緑体の分布は細胞の端っこ、つまり細胞膜に近い部分に限られます。さらに、柵状組織では細胞が円筒状に配置されているため、葉緑体は細胞壁に沿って縦に並ぶことになります(上の写真でもある程度わかります。細胞内部、周辺部分に並んでいる粒が葉緑体です。光の入射方向に対し平行に葉緑体が並んでいることになります。このように葉緑体の分布が不均一な状況では、葉緑体がランダムに分布しているときに比べると、葉緑体同士の相互被陰が大きくなり、個々の葉緑体が吸収する光の量が小さくなります。逆に、下部に漏れていく光の量は多くなります。葉緑体の分布が不均一であるために透過する光の量が多くなることをふるい効果(sieve effect)とか光チャネリング(light channeling)とか呼んでいるようです。相互被陰が大きくなるとか下部に漏れる光が多くなる、とか書いてしまうと不利な性質に思えますが、これは実は適応的な形態だと考えられています。上にも書きましたように、柵状組織はもともと光合成に利用しきれないような強い光があたるので吸収光率を高める必要がなく、海綿状組織に光を分配する効果があると言えます。
葉内の散乱の効果は特に緑の光で大きくなります。緑の光はクロロフィルが吸収しにくいので背軸側まで透過しやすいのですが、散乱によって吸収の確率が高まるわけです。クロロフィルを有機溶媒に溶かして全く散乱のない状態で吸光スペクトルを比べると、赤や青の吸収率が緑の吸収率に比べ5倍近く高いのですが、葉内で比べてみると、赤と緑の違いは20%程度になり、散乱によって緑の光が吸収されやすくなることがわかります。
藻類や植物では葉緑体が動くことが知られています。葉緑体は、紡錘型をしているイメージをお持ちの方も多いと思いますが、実際には扁平な円盤型をしています。光が強いときには葉緑体は細胞の側壁に並び、光を下部に通すように、光が弱いときには光をより受け取るように光に対して垂直に並びます。この意義はいうまでもなく、強すぎる光はできるだけ吸収せずに下部に逃がし、弱い光はできるだけ受け取る、というものです。
葉緑体運動の効果は葉の透過率を測定するとよくわかります。ムラサキツユクサでは、葉にあてる光強度が15μmol m-2 s-1くらいだともっとも多くの葉緑体が水平になります(柵状組織で40%、海綿状組織で70%)。光が強くなると半数以上の葉緑体が垂直になるのですが、葉の透過率は15μmol m-2 s-1で7%→飽和光で10%程度まで上昇します(Terashima and Hikosaka 1995。この実験には私は関与してませんが)。わずか数%ではありますが、状況に応じて光を逃がしたりより多く吸収するように工夫しているわけです。実際、葉緑体運動ができない突然変異体は光阻害を受けやすいようです(Kasahara et al. 2002)。また、葉緑体運動と透過率の関係を利用して、逆に透過率から野外での葉緑体運動の様子を探ろうとする研究もあるようです(Williams et al. 2003)。
高等植物では、葉緑体運動は細胞ごとにおこります。ムラサキツユクサでは、表から500μmol m-2 s-1の光をあてると、柵状組織では80%以上の葉緑体が垂直になるのに対し海綿状組織では半数程度の葉緑体しか垂直にはなりません。一方、裏から500μmol m-2 s-1の光をあてると、海綿状組織の葉緑体も70%までが垂直になります(Terashima and Hikosaka 1995)。
葉緑体運動については生理学的な面から多くの面で研究が進んでいます。例えばどのようなシグナル伝達が起こっているのかや、葉緑体が動くしくみなどですが、私には専門外なのでここではふれないでおきます。興味がある方は高木(2001)などをご覧下さい。
上で述べたように、葉内には向軸側が明るい、背軸側が暗いという光環境の勾配があります。葉の光合成特性が葉群内の光環境の勾配に応答して順化しているように、個々の葉緑体の光合成特性も光環境の勾配にあわせて順化しています。Terashima and Inoue (1984, 1985a) では、柵状組織と海綿状組織からそれぞれ葉緑体(チラコイド膜)を単離し、光合成特性を比較しました。柵状組織の葉緑体は最大電子伝達活性、光化学系I、IIの最大電子伝達活性が高く、さらに、チトクロムfやRuBPCaseなどのタンパク質も多く持っていました(クロロフィルあたり)。さらにTerashima and Inoue (1985b) では葉を10層に分けてチラコイド膜を単離し、chl a/b比や電子伝達活性が向軸側の葉緑体ほど高いことを明らかにしました。また、葉緑体の構造も異なり、グラナスタッキングの数やチラコイド膜/ストロマ体積比は背軸側の葉緑体ほど多くなっていました(Terashima et al. 1986, Terashima and Evans 1988)。これらの違いは陽葉・陰葉の違いと共通しています。陽葉・陰葉の違いについては「光順化」をご覧下さい。
このような葉緑体の性質にはある程度可塑性があります。Terashima et al. (1986) ではダイズの葉の裏を明るくし、表を暗くするような通常と逆の光環境で育てる、という実験をしました。展開が終了してから光環境を逆にする実験ではそれほど大きな変化が見られませんでしたが、展開途中の葉の光環境を逆にしてやると、それに合わせて葉緑体の構造などが変化するという現象が見られました。Evans et al. (1993) では、葉の角度を操作し、水平にした葉と垂直にした葉とで葉内の葉緑体の性質の分布が変化することが示されています。
このような葉緑体の性質の勾配は、個々の葉緑体の光合成の効率を上げるだけでなく、葉全体の光合成の効率も高めます。どういう理屈で高めるのかをこちらに書きました。
ここまでは葉内の光の分布の話でしたが、ここからはCO2拡散の話になります。拡散についてはこちらで説明しましたので一通り読んで下さい。
気孔から入ったCO2は細胞間隙を拡散し、葉肉細胞の表面(細胞壁)で水に溶け、細胞膜、細胞質、葉緑体包膜を経てストロマにたどり着きます。
光合成の話で重要なのは、これらの部位でCO2の拡散にどれだけの抵抗がかかるか、ということです。葉内、つまり気孔を通って以降の経路でかなりの抵抗がかかる、ということはここ10年あまりの研究で明らかになっています。条件の良いときならばCi/Ca比(細胞間隙と外気のCO2濃度比)とCc/Ci比(葉緑体と細胞間隙のCO2濃度比)は同レベルで(Evans and von Caemmerer 1991)、おおざっぱに言うと、気孔がもっとも開いているときと同じ程度の抵抗が葉内でかかります。詳しくはこちらに書きましたのでご覧下さい。
まずは細胞間隙について検討しましょう。かつてはCO2が細胞間隙を移動する間に相当の抵抗がかかるのではないかと考えられていました。厚い葉ほど光合成能力や窒素あたりの光合成速度が低いことが知られていましたが、その原因として、葉を厚くするとCO2が届きにくくなり、光合成にとってかなり不利になるのではないか、という議論がありました。今でもそういう考察をする論文もたまに見られます。しかし、細胞内のCO2拡散についての理論的研究の成果を見ると、細胞間隙内のCO2拡散抵抗は、葉肉細胞が密につまった下面気孔葉を除けば、それほど大きくないことがわかります(Parkhurst 1994, Terashima et al. 2001)。Parkhurst (1994) に示されている下面気孔葉についての計算例を見ると、気孔付近でCO2濃度の大きな低下(340ppm→230ppm)が起こりますが、そのあとは最上面でも205ppmまでしか落ちません。少なくとも気孔でのCO2拡散抵抗に比べればCO2濃度の低下はかなり小さいと考えていいようです。Evans et al. (1994) の実測でも葉内コンダクタンスと細胞間隙の長さ(葉の厚さ)とは相関がありませんでした。Evans and von Caemmerer (1996) によれば、両面気孔葉では葉内抵抗に対する気相での拡散の影響は10%程度でほとんど無視できるようです。ただし、細胞が密につまった下面気孔葉ではその影響は50%程度まで高くなると考えられているようです。
ということで、多くの葉では葉内コンダクタンスでは細胞壁で水に溶けて以降の経路が重要だと考えられています。実際、葉内コンダクタンスと「葉緑体が細胞間隙にさらされている面積を葉面積で割ったもの」(以下Sc)との間に高い相関があることがEvans et al. (1994) によって示されています。これはかいつまんでいうと、CO2が細胞表面でとけ込む面積が大きいほどコンダクタンスが高いことを意味し、細胞壁以降の経路が重要であることを強く示唆しています。細胞間隙と細胞壁以降では拡散経路の長さがずいぶん違いますが、水中でのCO2の拡散係数は空気中での10000分の1しかないので、短くとも影響は大きいのです。また、この経路を少しでも短くするために、葉緑体の分布は細胞の周辺部分に限られます。もし細胞周辺から離れてしまうと、光合成するためのCO2を受け取りにくくなることでしょう。
では細胞壁以降の経路の何がコンダクタンスに重要か、というのははっきりとはわかっていません。細胞壁は一つの候補です。葉肉細胞の細胞壁の厚さは種によって異なり、光合成能力が低い種ほど厚い傾向があるようです(Terashima et al. 1995)。このことから、細胞壁の厚さが葉内コンダクタンスに効いていることが示唆されます。もう一つは細胞膜のアクアポリンと呼ばれる膜タンパク質です。アクアポリンについて私はほとんど知識がありませんが、動物細胞ではアクアポリンが水やCO2を透過させるチャネルの役割をしていることが示されているようです。 Terashima and Ono (1992) は植物でもアクアポリンがCO2拡散に重要なのではないかと考え、アクアポリンの阻害剤であるHgCl2を葉に加え、葉緑体CO2濃度を測定し、HgCl2を加えた葉では葉緑体CO2濃度が大きく低下していることを見出しました。このことからアクアポリンがCO2拡散に関与していることは示唆されますが、実際の葉でどれだけの抵抗を持っているかなど、まだ解明すべき点は多いようです。このあたり、寺島(2003)もご覧下さい。
以上をふまえ、光合成速度を上げるために葉がどのようにあるべきか、というのを考えてみましょう。
まず、光合成能力(飽和光下の光合成速度)を高くするためには、RuBPCaseなどの光合成系タンパク質を多く持つ必要があります。光合成系タンパク質を保持するためには、意外に思われるかもしれませんが葉緑体の体積が重要です。RuBPCaseなどカルビンサイクルの酵素はストロマ中に溶けていますが、その濃度は飽和に近いと考えられています(Bjorkman 1981)。実際、葉内の葉緑体の体積と光合成能力は高い相関があります(Ono et al. 1995, Oguchi et al. 2003)。したがって、光合成能力を増やすためには、葉緑体の数を増やすか大きくするかのどちらかが必要です。
葉緑体のの形や位置も重要です。葉緑体が扁平であると上に書きましたが、これには理由があります。細胞間隙からCO2を受け取るためです。まず、葉緑体は細胞膜に近い部分に配置しなければいけません。次に、葉緑体の厚さを薄くしなければいけません。水中ではCO2の拡散は非常に遅いですから、葉緑体が細胞膜から離れていたり、球体だったりすると、細胞膜から離れている部分のRuBPCaseはCO2を受け取ることができません(Terashima et al. 2001、 寺島 2003、後者では寺島さんのマンガを見ることができます)。
これらのことから、光合成能力を増やすには葉緑体数もしくは体積を増やすだけでは不充分で、葉肉細胞の表面付近に葉緑体を並べるための面積が必要であることがわかります。葉肉細胞の表面積を増やす方法はいくつか考えられますが、実際に葉が行っていることは一つ、「葉を厚くすること」です。これが陽葉が陰葉よりも厚い理由です。陽葉は葉緑体をたくさん収容するために葉を厚くし、陰葉はその必要がないから薄い葉を作るわけです。暗い環境では光が不足しますから、同じ量のバイオマスをもっているならば、面積が小さい厚い葉を作るよりは薄くて広い葉を作ったほうが得です。
ここで一つ面白い話をしましょう。陰葉は厚さが薄いわけですが、陰葉を強光に移すと光合成能力が上がることが多くの種で知られています。一方葉の形態は、一部の種を除くと、成熟したあとには細胞の形などはほとんど変化しないといわれています。葉が薄いままどうやって光合成能力を増加させているのでしょうか? この疑問を実際に調べてみたのがうちの小口くんです。彼は一年草シロザを弱光で育て、葉の展開が終了したのち強光に移しました。光合成能力は約1.5倍増加しました。葉の内部を顕微鏡観察し、解析してみると、光合成能力は「葉緑体が細胞表面に接している面積(以下Sc)」と高い相関があることがわかりました。強光に移した葉ではScが増加したわけですが、どのようにして増加が可能になったのでしょうか? それは、弱光生育の葉にあらかじめ「すきま」があったからでした。細胞表面には葉緑体がないスペースがあり、強光に移すと葉緑体の面積が広がり、このスペースが埋まったのです。つまり弱光に育っている葉は強光になることを期待し、あらかじめスペースを空け、葉緑体を増やす余地を残していたと考えられます。詳しくはOguchi et al. (2003) をご覧下さい。