光合成の生理生態学講座
光合成の温度依存性の性質 (200601 13)
弱光下の光合成速度 (200601 13)
強光・低CO2濃度での光合成速度 (200601 13)
強光・高CO2濃度での光合成速度 (200601 24)
リン酸律速時の光合成速度 (200601 24)
結局光合成速度の温度依存性はどうなるのか? (200601 24)
多くの場合酵素の反応速度は温度の影響を強く受けます。反応速度は温度上昇とともに増加し、あまり高くなると酵素が失活して速度が下がってしまう、というのが一般的です。光合成も温度の影響を受けますが、その影響はこのような単純なものではありません。
極端な低温・高温は光合成系にダメージを与えます。しかしこの影響も様々な種類があり、私自身はあまり知識がないので、この項には書きません。低温が引き起こすダメージの一つは光化学系における光阻害です(低温下では光阻害がかかりやすい)。光阻害については別に書きます。
この項で扱うのは、可逆的な光合成速度の変化、つまり光合成系にダメージを与えない程度の温度変化です。また、この項での説明は基本的にFarquharのモデルにしたがったものになります。Farquharのモデルをおおざっぱにでも理解していなければ本項のみならず、光合成速度の温度依存性を理解することは難しいと考えていいと思います。数式まで理解する必要はありませんが、「二酸化炭素と光合成」の「直観的な説明」には目を通しておいて下さい。
詳しい総説としては、古くはBerry and Bjorkman (1980)、最近では私のレビュー(Hikosaka et al. 2006)をご紹介しておきます。
光合成速度の温度依存性(いわゆる温度-光合成曲線)というと、山型(ベル型)のカーブを思い浮かべると思います。これは一般的に通常CO2濃度、飽和光強度で測定したものです。測定条件が変わるとカーブの形が変わります。
下の図に示したのは異なるCO2濃度で測定した温度-光合成曲線です(強光下)。見ての通り、CO2濃度によって速度が最大になる温度(最適温度)が異なり、高CO2濃度ほど最適温度が高くなります。
白丸が1000ppm、黒丸が370ppm(通常CO2濃度)、四角が200ppm。オオバコのデータ。測定は私。
光条件によっても異なります。あいにく手持ちのデータがないのですが、弱光で測定するほど最適温度が低くなります。光補償点付近では、温度-光合成曲線はベル型ではなく低温ほど高くなります。
このように条件によって温度依存性が変化するのは、律速段階によって反応速度の温度依存性が異なるためです。まず、光化学反応は、ほとんど温度依存性を示しません(温度依存性があるのかもしれないが、ものすごく短い時間で起こるので無視できる?)。しかし、他の反応のほとんど-強光下の電子伝達速度、ATPase合成速度、カルビンサイクルの各反応-は温度依存性を示します。非常に重要なことは、RuBPCaseが触媒する二つの反応-RuBPのカルボキシル化と酸素化-は温度依存性が異なる、ということです。どちらの反応も温度が高くなると活性が上がる、ということには違いがないのですが、おおざっぱに言って酸素化反応速度のほうが上がり方が急です。したがって、光合成と光呼吸を相対的に比べると、高温ほど光呼吸のほうが高い、ということになります。
以下の項では、なぜ測定条件によってカーブの形が変わるのかを通して温度-光合成曲線の成り立ちを説明します。
弱光下の光合成速度は暗呼吸速度と初期勾配の影響を受けます。
暗呼吸速度は一般に温度が高いほど高くなります。弱光下で光合成速度が低下する理由の一つがこれです。暗呼吸については私はほとんど知識がありません。Lloyd & Taylor (1994) は土壌呼吸の温度依存性をいろんな文献から探してきて、どういう曲線が一番フィットするか、というのをやっていました。だいたい指数関数に近い曲線で回帰できるようですが、彼らは最終的にずいぶん凝った式で回帰していました。以下に示すのはシラカシ(Quercus myrsinaefolia)の葉で得られたカーブです。温度依存性は生育条件に依存しません。まあだいたい指数関数的ではないかと思います。
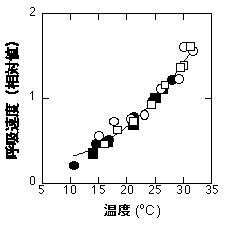
シラカシの呼吸速度の温度依存性。白丸は30度で育った葉の、黒丸は15度で育った葉のデータ。25度での速度を1としている。測定は村上綾子氏。
次に初期勾配ですが、上述のように、光が律速しているときの光化学反応の速度は温度に依存しません。このためRuBPの生産速度も温度に依存しませんが、RuBPが酸素化に利用されるかカルボキシル化に利用されるかの割合が温度によって変化します。上に述べたように、酸素化反応は相対的に高温で起こりやすいため、大気CO2濃度条件では高温ほど初期勾配が低下します(Ehleringer & Bjorkman 1977)。これも高温で光合成速度が低下する理由の一つです(ただしその影響は呼吸に比べると小さい)。しかし、かなり高いCO2濃度条件もしくは低酸素条件では光呼吸が起こらないので、初期勾配は温度によって変化しません。ちなみに、C4植物では光呼吸が起きないので、初期勾配の温度依存性はありません。
こちらで説明したとおり、RuBPが飽和しているときにはRuBPCaseのキネティクスで光合成速度が決まります(Vcmax・Kcについてはこちらをご覧下さい)。
まず基本的なRuBPCase活性の温度依存性について記します。まずカルボキシル化について考えるために、仮にO2濃度をゼロとし、酸素化が起こらないと仮定しましょう。CO2濃度が飽和しているときには、カルボキシル化速度(つまりVcmax)は温度に対して指数関数的に増加します(Jordan and Ogren 1984)。一方、Kc(CO2に対するRuBPCaseの親和性)も指数関数的に増加します。Kcが増加するということは親和性が低下するということで、速度を低下させる方向にはたらきます。低CO2濃度では、温度増加に対するVcmaxの増加とKcの増加がうち消し合います。Vcmaxの影響は低温で大きく、Kcの影響が高温ほど大きいため、低温では温度上昇とともにカルボキシル化速度が増加し、高温では低下します。この結果カーブの形が山型(ベル型)になります。低CO2濃度ほどKcの効果が大きいため最適温度が下がります。
通常大気条件ではさらにこれに光呼吸の影響が加わります。上記のように、酸素化の温度依存性はカルボキシル化の温度依存性よりも大きいため、温度が高ければ高いほど光呼吸によるCO2放出が大きくなります(こちらの式8で言うと、G*が増加する)。このためますます光合成速度は高温で低下するようになり、最適温度が低くなります。光呼吸の影響は高CO2で小さくなりますので、高CO2ほど最適温度が高くなります。
これらの結果、RuBPCaseが律速しているときの光合成速度 (Pc)と温度の関係はベル型となり、最適温度はCO2濃度が高いほど高くなります。ただし、高CO2では普通RuBP再生が律速するので、現実の光合成速度の温度依存性はまた別の話になります。
以下の図は、RuBP濃度が飽和しているときの光合成速度です。Vcmax、つまり飽和CO2濃度でのカルボキシル化速度の温度依存性)と通常CO2濃度(ベル型)での光合成速度(Vcmaxの温度依存性から推定したもの)を示しています。
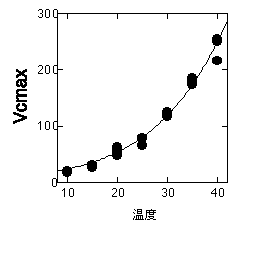
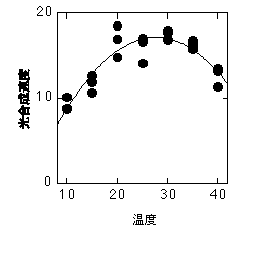
Vcmaxの温度依存性は、オオバコから得られたもの。石川数正氏測定。
この図ではVcmaxは指数関数的に増加していますが、種によっては高温で指数関数から外れたりVcmaxが下がったりすることがあります。
強光・高CO2濃度での光合成速度(RuBPが律速しているときの光合成速度)
こちらに書いたように、Farquharらのモデルでは、RuBP供給速度を律速しているのは電子伝達系であると仮定してRuBPが律速しているときの光合成速度をモデル化しています。光飽和時には最大電子伝達速度(Jmax)とG*のみが光合成速度に影響します。上述のようにG*は高温ほど高くなります。Jmaxの温度依存性は多くの場合30-40度付近でピークがあるカーブになります。
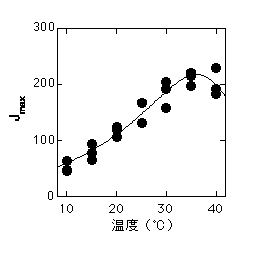
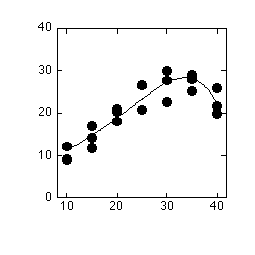
左はオオバコのJmaxの温度依存性。右はそのJmaxの温度依存性から計算した、大気CO2濃度での光合成速度。最適温度がちょっとだけ低温にずれる。Jmaxは石川数正氏測定。
Jmaxの温度依存性の形というのは、ほぼそのまま飽和CO2濃度での光合成速度の温度依存性の形と同じものになります(リン酸律速が起こっていない場合)。低CO2では、G*の影響で高温ほど光合成速度が低めになり、この結果最適温度が若干低温にずれます。
トリオースリン酸の利用速度は非常に温度依存性が大きいようです。Harley et al. (1992) やCen and Sage (2005) に測定例が示されています。トリオースリン酸が律速している場合には、光合成速度はCO2濃度・O2濃度・光強度に依存しませんので、トリオースリン酸の利用速度の温度依存性と光合成速度の温度依存性は同じ形になります。
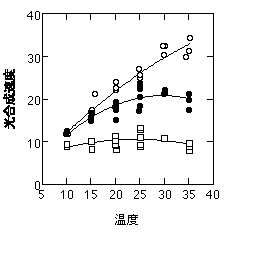
ここまで各部分反応の温度依存性について書いてきましたが、どの部分反応が光合成速度を律速するかは種や生育温度によって異なります。多くの場合光合成速度はRuBPCaseに律速されますが(Pc)、シラカシの場合、30度で育ててると、大気CO2濃度での光合成速度は23度より上でRuBPCaseに、23度より下ではRuBP再生に律速されることが示されました(Hikosaka et al. 1999)。
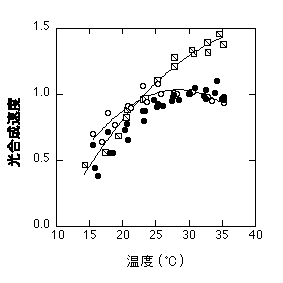
シラカシの温度-光合成曲線。黒丸が実測した光合成速度(通常CO2濃度・飽和光)。白丸は低CO2濃度の光合成速度から推定した、通常CO2濃度でのPc。四角は高CO2濃度での光合成速度から推定した、通常CO2濃度でのPr。光合成速度は低温でPrに、高温でPcに等しいことがわかる。
各部分反応の温度依存性にも種間差・生育条件による違いがあり、これらの結果、温度-光合成曲線の形は生育条件や種によって異なる、ということになります。生育条件による違いは「温度順化」で説明します。
一つ大事なことを書いておきます。例えばCO2濃度が変わると温度依存性が変化し、最適温度が変化します。一見するとカーブが水平に移動したかのような印象を受けますが、生化学的には違います。温度によって光合成速度のCO2濃度依存性が異なるために見かけ上最適温度が動くのです。高CO2濃度での最適温度における光合成の性質(例えば酵素の活性)と低CO2濃度での最適温度における性質には直接的な関係はありません。