岝崌惉偺惗棟惗懺妛島嵗
丂
偼偠傔偵丂
岝壔妛宯II丂(200510 05丒200510 10乯
岝壔妛宯I丂(200510 10乯
丂
乽岝崌惉偺婡嶌乿偱偼岝崌惉宯慡懱偐傜橂嵴偟偨岝崌惉偺偟偔傒傪愢柧偟傑偟偨丅偙偙偱偼丄奺斀墳傪扴偆岝崌惉宯峔惉梫慺偵偮偄偰嵶偐偄夝愢傪偟傑偡丅慡偰偺峔惉梫慺偵偮偄偰愢柧偡傞偺偱偼側偔丄乮懠偺晹暘偺乯昁梫偵墳偠偰弴師愢柧傪壛偊偰偄偔偙偲偵側傞偐偲巚偄傑偡丅
丂岝壔妛宯II偼僠儔僐僀僪忋偵晜偐傇僋儘儘僼傿儖偲僞儞僷僋幙偺暋崌懱偱丄20庬埲忋偺僒僽儐僯僢僩偐傜峔惉偝傟偰偄傑偡丅岝壔妛宯II暋崌懱偼丄戝偒偔僐傾偲傾儞僥僫偵暘偗傞偙偲偑偱偒傑偡丅僐傾偱偼丄壓偵婰偡傛偆偵斀墳拞怱偲揹巕揱払峔惉惉暘偑偁傝丄揹巕揱払偑婲偙傝傑偡丅傾儞僥僫偼廤岝惈僋儘儘僼傿儖亅僞儞僷僋幙暋崌懱乮LHCII, Light harvesting chlorophyll-protein complex乯偲傕屇偽傟丄岝傪曔廤偡傞栶妱傪帩偪傑偡丅
丂LHCII偼丄堦偮偺僒僽儐僯僢僩乮億儕儁僾僠僪乯偑12偐13偺僋儘儘僼傿儖傪寢崌偟偰偍傝乮Kuhlbrandt et al. 1994乯丄暘巕検偑25kD慜屻偱偡丅崅摍怉暔偺傾儞僥僫偼懡偔偺応崌擇庬椶偺LHCII僒僽儐僯僢僩乮Lhcb1, Lhcb2乯偐傜側傞傛偆偱偡乮Jansson 1994乯丅堦偮偺僐傾偵懡偔偺LHCII偑寢崌偟丄暥帤捠傝乽傾儞僥僫乿偲側偭偰岝偺曔廤傪偟傑偡丅傾儞僥僫偵寢崌偟偰偄傞僋儘儘僼傿儖偼丄岝傪媧廂偡傞偲椼婲偝傟傑偡乮excitation乯丅僋儘儘僼傿儖偑媧廂偟偨僄僱儖僊乕偼椼婲僄僱儖僊乕偲偟偰師乆偵暿偺僋儘儘僼傿儖偵揱払偝傟丄嵟廔揑偵丄僐傾偵懚嵼偡傞斀墳拞怱P680偵揱払偝傟傑偡丅偙偺P680偑椼婲僄僱儖僊乕傪庴偗庢傞偙偲偱揹巕揱払偑巒傑傝傑偡丅
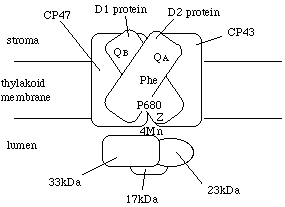
丂僐傾偱偼揹巕揱払偑婲偙傝傑偡丅壓恾偼崅摍怉暔偺岝壔妛宯II僐傾偺娙扨側柾幃恾偱偡丅偙偙偱偼丄偐側傝娙棯壔偟偰偁傞偺偲丄LHCII偼徣偄偰偁傞偙偲偵拲堄偟偰壓偝偄丅僐傾偺拞怱晹暘偼D1僞儞僷僋幙偲D2僞儞僷僋幙偺擇検懱偱偡丅廃埻偵CP47傗CP43傪偼偠傔懡偔偺僒僽儐僯僢僩偑偁傝傑偡丅僐傾偩偗偱栺400-500kDa偵側傝傑偡丅
丂
丂
丂揹巕偺棳傟傪扨弮偵彂偔偲丄悈仺儅儞僈儞僋儔僗僞乮恾偱偼4Mn乯仺Z仺P680仺僼僃僆僼傿僠儞乮Phe乯仺QA仺QB偲側傝傑偡丅偨偩丄悈暘夝偐傜揹巕揱払偑巒傑傞傢偗偱偼偁傝傑偣傫丅
丂P680偼埫崟壓偱偼丄娨尦偝傟偨忬懺偵偁傝傑偡乮揹巕傪帩偭偰偄傞乯丅P680偼丄僄僱儖僊乕傪庴偗庢偭偰椼婲偝傟傞偲丄僼僃僆僼傿僠儞偵揹巕傪搉偟傑偡丅僼僃僆僼傿僠儞偐傜偼揹巕偼QA丄QB偲搉傝傑偡丅P680偼僋儘儘僼傿儖a偺擇検懱偲峫偊傜傟偰偄傑偡丅僼僃僆僼傿僠儞偲偄偆偺偼丅僋儘儘僼傿儖a偐傜儅僌僱僔僂儉僀僆儞偑奜傟偨傕偺偱偡丅岝壔妛宯II偵偼俀暘巕懚嵼偟傑偡偑丄揹巕揱払偵偼偨傜偄偰偄傞偺偼堦暘巕偩偗偺傛偆偱偡丅QA丆QB偼偳偪傜傕寢崌宆偺僾儔僗僩僉僲儞偱偡丅QB晹埵偱揹巕傪庴偗庢偭偨僾儔僗僩僉僲儞偑梀棧偟偰cyt b/f暋崌懱偵揹巕傪揱払偟傑偡丅P680丄僼僃僆僼傿僠儞丄QB偼D1僞儞僷僋幙偵丄QA偼D2僞儞僷僋幙偵寢崌偟偰偄傑偡丅
丂偝偰丄岝僄僱儖僊乕偵傛傝椼婲偝傟丄巁壔偝傟偨乮揹巕傪搉偟偰偟傑偭偨乯P680偵偼揹巕偑曗廩偝傟傑偡丅偙偺揹巕偼悈傪暘夝偡傞偙偲偱摼傜傟傑偡丅悈偼儅儞僈儞僋儔僗僞偱暘夝偝傟丄庢傝弌偝傟偨揹巕偼Z傪宱偰P680偵搉偝傟傑偡丅儅儞僈儞僋儔僗僞偼4偮偺儅儞僈儞尨巕偐傜側傝傑偡丅悈傪暘夝偡傞儊僇僯僘儉偼枹偩偵柧傜偐偱偼側偄傛偆偱偡偑丄偐側傝妋搙偑崅偄儌僨儖偑弌偝傟偰偄傑偡丅悈傪暘夝偟偰巁慺偑弌傞傑偱偵偼丄係岝検巕偑岝壔妛宯II偵摉偨傞偙偲偑昁梫偱偡丅P680偑巁壔偝傟傞偨傃偵儅儞僈儞僋儔僗僞偑揹巕傪搉偟偰曄宍偟乮S0丄S1丒丒丒S4偲屇偽傟傞乯丄4夞揹巕傪搉偡偛偲偵悈傪暘夝偟丄巁慺傪敪惗偝偣偰偄傞偺偩傠偆偲峫偊傜傟偰偄傑偡丅Z偼丄D1僞儞僷僋幙偺僠儘僔儞巆婎偱偡丅岝崌惉偺揹巕揱払宯偱傾儈僲巁偑捈愙揹巕揱払偺栶妱傪偡傞偺偼Z偩偗偱偡丅
丂岝壔妛宯II偑帩偮僋儘儘僼傿儖偺悢偼惗堢忦審偵傛偭偰戝偒偔曄傢傝傑偡丅偙傟偼丄惗堢忦審偵傛偭偰堦偮偺僐傾偵寢崌偡傞LHCII偺悢偑曄傢傞偨傔偱偡丅LHCII偺悢偺曄壔偼僋儘儘僼傿儖a偲僋儘儘僼傿儖b偺斾傪尒傞偙偲偱斾妑揑娙曋偵抦傞偙偲偑偱偒傑偡丅僐傾偵寢崌偟偰偄傞僋儘儘僼傿儖偺傎偲傫偳偼僋儘儘僼傿儖a偱偡偑丄LHCII偼僋儘儘僼傿儖a偲僋儘儘僼傿儖b傪敿乆偔傜偄偯偮帩偭偰偄傞偨傔丄LHCII偑憹偊傞偲僋儘儘僼傿儖a/b斾偑尭彮偟傑偡丅
丂岝壔妛宯II偺僐傾偵偼CP47偲CP43偲偄偆僋儘儘僼傿儖傪寢崌偟偰偄傞僒僽儐僯僢僩偑偁傝傑偡乮"CP"偼偨傇傫chlorophyll Protein丄悢帤偼揹婥塲摦偟偨偲偒偺尒偐偗偺暘巕検kD乯丅偙偺懠丄CP29乮Lhcb4乯丄CP26乮Lhcb5?乯丄CP24乮Lhcb6乯偲偄偆僒僽儐僯僢僩傕偁傝傑偡丅偙偺3偮偺僒僽儐僯僢僩偼LHCII偺僒僽儐僯僢僩乮Lhcb1, Lhcb2乯偵傛偔帡偰偍傝丄婡擻揑偵偼傾儞僥僫偲峫偊傞偺偑懨摉偱偡偑丄堦偮偺僐傾偵偍偦傜偔堦僒僽儐僯僢僩偟偐側偔丄惗堢忦審偵傛傞検偺曄壔偼側偄傛偆偱偡乮Bailey et al. 2001乯丅巹偑岝崌惉偺拏慺暘攝偺儌僨儖傪嶌偭偨偲偒乮Hikosaka and Terashima 1995乯偵偼丄堦偮偺LHCII僒僽儐僯僢僩偑13丄僐傾偑60帩偭偰偄傞偲壖掕偟傑偟偨丅岝壔妛宯II暋崌懱偲偟偰偼100-300掱搙偺僋儘儘僼傿儖傪帩偭偰偄傞偙偲偵側傞偼偢偱偡丅
丂
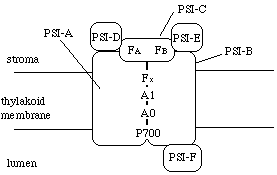
丂岝壔妛宯I偺峔憿丄婡擻偼岝壔妛宯II偵傛偔帡偰偄傑偡丅暋崌懱偼傗偼傝悢廫庬偺僒僽儐僯僢僩偐傜峔惉偝傟丄傾儞僥僫乮LHCI乯傪帩偭偰偄傑偡丅LHCI偺僋儘儘僼傿儖偑岝傪媧廂偡傞偲丄斀墳拞怱P700偵椼婲僄僱儖僊乕偑揱払偝傟傑偡丅揹巕偼丄P700仺A0仺A1仺Fx仺FA/FB偲偄偆弴偵揱払偝傟丄僼僃儗僪僉僔儞偵搉偝傟傑偡丅丅巁壔偝傟偨P700偵偼僾儔僗僩僔傾僯儞偑捈愙揹巕傪搉偟傑偡丅
丂700偼僋儘儘僼傿儖a偺擇検懱偲峫偊傜傟偰偄傑偡丅A0傕僋儘儘僼傿儖偲峫偊傜傟偰偄傑偡丅偨偩偟丄偙偪傜偼堦暘巕偱偡丅A1偼擇暘巕偺價僞儈儞k1偲峫偊傜傟偰偄傑偡丅Fx丄FA/FB偼偲傕偵揝亅棸墿僙儞僞乕偱偡丅
丂僒僽儐僯僢僩偺峔憿傕傛偔帡偰偄傑偡丅拞怱晹偵偼PSI-A偲PSI-B偲偄偆僒僽儐僯僢僩偑擇検懱傪宍惉偟偰偄傑偡乮岝壔妛宯II偺D1丄D2偵憡摉乯丅偙偺擇検懱偵P700偑寢崌偟偰偄傑偡丅PSI-C偼擇偮偺揝亅棸墿僙儞僞乕傪帩偭偰偄偰丄揹巕揱払偵娭學偟傑偡丅PSI-D偲PSI-E偼僗僩儘儅懁偵埵抲偟丄僼僃儗僪僉僔儞偲岝壔妛宯I偺揹巕揱払傪庤揱偄傑偡丅摨條偵丄PSI-F偼僾儔僗僩僔傾僯儞偲岝壔妛宯I偺揹巕揱払傪庤揱偄傑偡丅埲忋丄Golbeck (1992) 偺廍偄撉傒偱偟偨丅
丂岝壔妛宯I偐傜揹巕傪庴偗庢偭偨僼僃儗僪僉僔儞偼丄僼僃儗僪僉僔儞-NADP巁壔娨尦峺慺傪夘偟偰NADP傪娨尦偟傑偡丅偙偆偟偰偱偒偨NADPH偑娨尦椡偲偟偰棙梡偝傟傞偙偲偵側傝傑偡丅僼僃儗僪僉僔儞偺婡擻偼偙傟偩偗偱偼側偔丄岝屇媧丄垷徤巁娨尦丄僇儖價儞僒僀僋儖偺偄偔偮偐偺峺慺偺妶惈壔偱偼捈愙娨尦椡傪嫙媼偟傑偡丅
丂岝壔妛宯I偺僐傾偲LHCI偺検斾偼丄岝壔妛宯II偵斾傋傟偽偁傑傝曄傢傜側偄傛偆偱偡丅扨棧偟偨岝壔妛宯I偼斀墳拞怱偁偨傝栺200偺僋儘儘僼傿儖傪帩偮偙偲偑懡偄傛偆偱偡丅巹偺儌僨儖偱偼斀墳拞怱偁偨傝184屄偱屌掕偟傑偟偨乮Hikosaka and Terashima 1995乯丅傑偨丄岝壔妛宯I暋崌懱偺暘巕検偼僐傾+LHCI偱栺500kDa偵側傝傑偡丅
丂