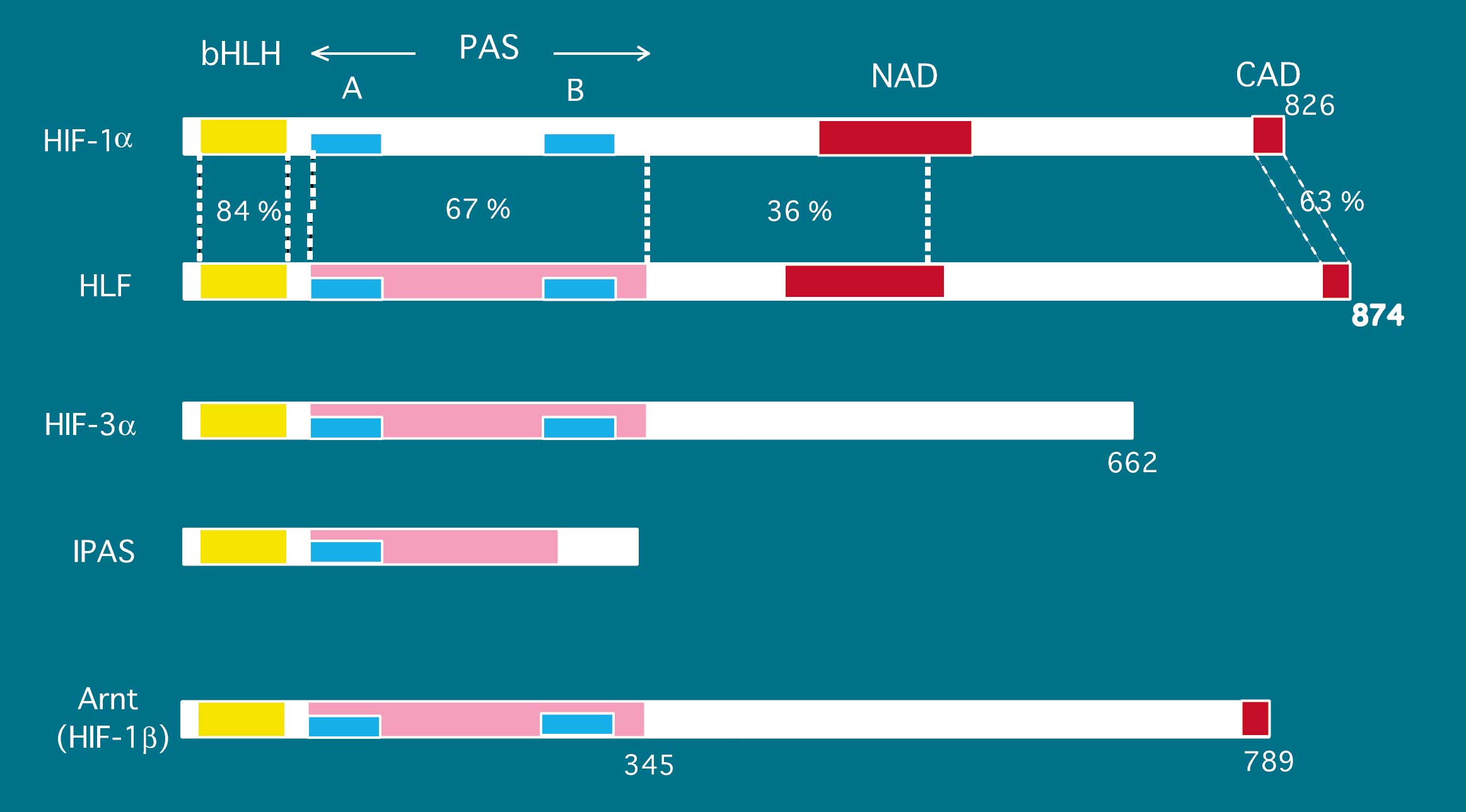
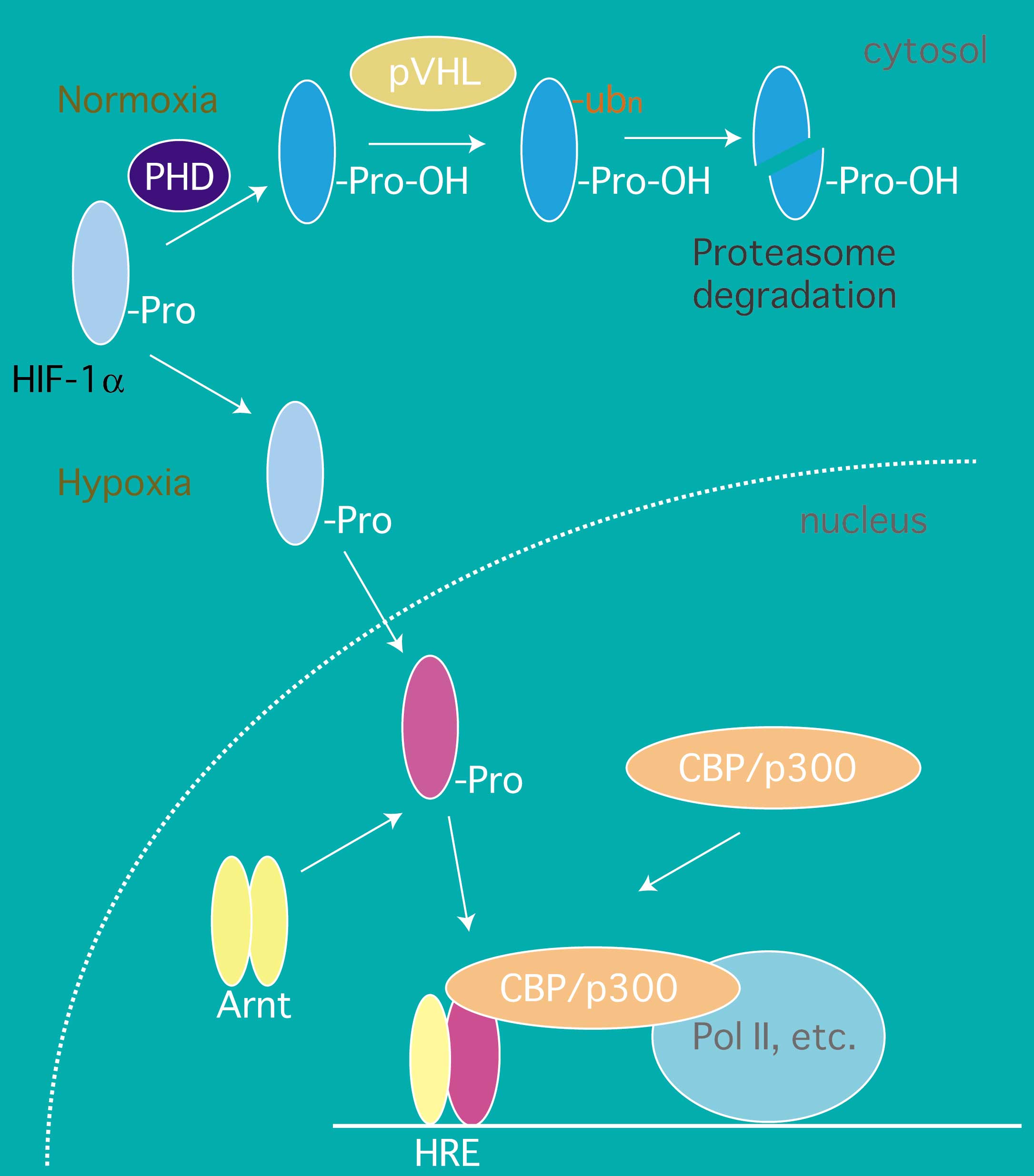
低酸素状態のことをハイポキシアといいます。ハイポキシアでに活性化される転写因子、Hypoxia-Inducible Factor-1(HIF-1)が見いだされて、低酸素応答系の分子機構の研究は急速に進展しました。また、類似因子HLFも見出され、高等動物でのハイポキシアに応答する遺伝子調節機構もかなり解明されてきています。