光合成の生理生態学講座
目次
はじめに (200301 26)
呼吸とは (200301 26)
解糖系・ペントースリン酸経路 (200301 26)
TCAサイクル (200301 31)
電子伝達系一般 (200302 17・20)
植物ミトコンドリアの電子伝達系 (200303 01)
植物も動物と同じように呼吸をします。光呼吸(photorespiration)とは全く別の経路なので、暗呼吸(dark respiration)と区別されることもあります。この「光合成の生理生態学講座」は文字通り光合成のためのページで、また、光合成と呼吸は全く違うプロセスなのですが、炭素の出入りという点からすると非常に重要なので、このページにも書くことにします。
私は呼吸に関しては素人なので、ここに書くことは主に教科書(「代謝」山谷編 2001、「植物生化学」Heldt 1999 [金井訳 2000]、Plant Physiological Ecology Lambers et al. 1998)に基づいています。もちろん責任は私にあります。
呼吸とは、糖などの有機物を分解してエネルギーを取り出す経路です。高校の教科書に載っているような式では、
C6H12O6 + O2 → CO2 +H2O + エネルギー
で、C6H12O6 に対し38ATPが生産される、とか教えていたと思います(今どうなっているか知りませんが)。この「38ATP」は「必ずこれだけ生産される」という数字ではなく、「上限」くらいに考えればいいと思います。
おそらく植物細胞中で起こっている呼吸の主要なものは上記経路で表されるものと同じだと思いますが、呼吸は糖だけではなく脂質やアミノ酸などを基質とすることができ、多様なバリエーションがあるのだと考えていいと思います。
呼吸の大部分はミトコンドリアで行われます。ミトコンドリアは細胞内小器官の一種で、多くの場合葉緑体よりかなり小さいです。外膜・内膜の二つの膜構造からなります。内膜の内側をマトリックスと呼びます。
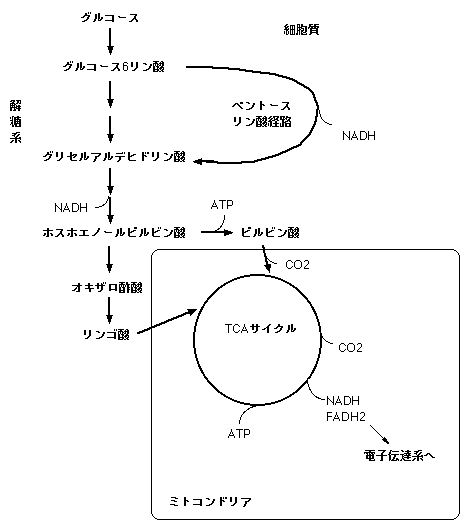
まずは呼吸の代謝経路を順を追って説明しましょう。呼吸は大きく三つのステップに分けることができます。第一のステップが細胞質で起こる解糖系(glycolysis)です。教科書的に解糖系を一言で説明すると、「糖を分解して有機酸(ピルビン酸)を合成し、ATPを取り出す」ということになります。ただしこれは動物細胞の場合で、植物細胞の場合、最終産物はピルビン酸ではなくリンゴ酸であることが多いそうです(少なくともLambers et al. 1998はそう書いています)。最終産物がリンゴ酸の場合は、解糖系ではATPは合成されません。
解糖系の最初のステップは、糖の分解です。ソースがグルコース(ブドウ糖)かスクロース(ショ糖)で若干経路が違いますが、まずグルコース6リン酸が作られます。ここまでの時点でATPを消費します。このあと複雑なステップを経て、ホスホエノールピルビン酸(PEP:C4光合成でCO2 固定に使われる物質と同じ)が合成されます。ここまでの間にATPが合成され、ATPの収支は0になります。このあとピルビン酸キナーゼが触媒してピルビン酸が合成される場合、ATPが生産されます。解糖系で生産されるエネルギーはこれだけです。リンゴ酸が最終産物になる場合は、PEPからオキザロ酢酸が合成され、さらにリンゴ酸が生産されます。ピルビン酸、リンゴ酸ともミトコンドリアに輸送され、TCAサイクルに入ります。
嫌気状態ではミトコンドリアで呼吸が行われなくなるため、ピルビン酸はミトコンドリアに入らず、乳酸発酵やアルコール発酵の経路に入ります。
細胞質には解糖系の他にもう一つグルコースを分解する系があり、ペントースリン酸経路(pentose phosphate pathway)と呼ばれています。ペントースリン酸経路では、最初のステップはグルコース6リン酸の分解です。複雑なステップを経てグリセルアルデヒド3リン酸(GAP)が生じます。GAPは解糖系の中間代謝産物でもあり、ここから解糖系に合流します。ペントースリン酸経路の役割は、五単糖リン酸とNADPHの供給にあると考えられています。ペントースリン酸経路の中間代謝産物には五単糖リン酸の一種であるリブロース5リン酸があり、これはリボースやデオキシリボースなどの拡散生合成の出発点になります。また、エリスロース4リン酸はシキミ酸経路に入り、フェノール化合物の生産に利用されます。NADPHは脂肪酸の生合成などに利用されたり、ミトコンドリアに輸送されて電子伝達に利用されたりします。ペントースリン酸回路はどちらかというと生体成分生合成における役割が大きく、細胞内のエネルギー収支に対する寄与は解糖系ほど大きくはないと思われます。実際、ペントースリン酸回路がエネルギー要求に応じてコントロールされているという証拠はないそうです(Lambers et al. 1998)。
解糖系で生産されたピルビン酸もしくはリンゴ酸はミトコンドリアに輸送され、TCAサイクル(クレブス回路・クエン酸回路とも)に入ります。TCAサイクルの役割は、ピルビン酸やリンゴ酸を分解して還元力(NADH・FADH2)を取り出すことにあります。また、ATPもわずかですが生産されます。このサイクルでピルビン酸は分解され、CO2が放出されます。呼吸で放出されるCO2はこれに由来します。
以下は細かい話です。TCAサイクルでは、ピルビン酸が輸送された場合は、まずピルビン酸とCoA(補酵素A)が結合し、CO2とアセチルCoAが生産されます。アセチルCoAとオキザロ酢酸が結合してクエン酸(とCoA)ができます。このクエン酸から再びオキザロ酢酸が生産され、サイクルとなります。このサイクルの間に、(1グルコースあたり)8NADHと2FADH2(コハク酸)の還元力と2ATPが生産されます。TCAサイクルにおいてオキザロ酢酸の一つ前のステップはリンゴ酸です。解糖系でリンゴ酸が生産された場合は、ここからTCAサイクルに入ることになります。
ミトコンドリアの電子伝達系の主な目的はミトコンドリア内膜の内外にプロトン勾配を作ることです。どうやってプロトン勾配を作るかというと、NADHなど還元力が強い物質から電子が伝達され、内膜中を移動する間にプロトンを膜の内側から外側に移動させるわけです。
では細かい話です。動物のミトコンドリアの電子伝達系には4つの大きな複合体があります。複合体I、II、III、IVと略称されます。複合体Iの名称は、NADHデヒドロゲナーゼです。これがNADHから電子を受け取り、ユビキノンと呼ばれるキノンに電子を渡します。その間にプロトンを膜の内側から外側に輸送します。ユビキノンは、(プラストキノン同様)膜内を移動し、複合体IIIであるチトクロムb/c複合体に電子を伝達します。その際、(やはりプラストキノン同様)プロトンを膜の内側から外側に運搬します。ユビキノンに電子を渡す複合体はもう一つあり(複合体II)、コハク酸デヒドロゲナーゼといいます。これはコハク酸(FADH2)から電子を受け取り、ユビキノンに電子を渡します。コハク酸デヒドロゲナーゼはプロトン輸送は行いません。
チロクロムb/cからはチトクロムcと呼ばれるタンパク質が電子を受け取ります。チトクロムcは膜中ではなく内膜の外側を移動し、複合体IVのチトクロムオキシダーゼに電子を伝達します。チトクロムオキシダーゼは酸素に電子を伝達し、水を作ります。この際にプロトンが輸送されます。
このようにしてプロトンの勾配が形成されると、その勾配をエネルギー源としてATP合成酵素によりATPが合成されます。原理は葉緑体でのATP合成とほとんど同じです。
「電子伝達系」というのは葉緑体のチラコイド膜にもあります。電子を伝達する系であること、ATPを合成するために膜内外にプロトン勾配を形成することなどが同じですが、電子の流れ方に大きな違いがあります。チラコイド膜の場合は、電子は還元力が弱い(=酸化還元電位が高い)水から還元力が強いNADPに電子が渡るのに対し、ミトコンドリア内膜では還元力の強さに依存して電子が移動します。この違いは前者で光エネルギーが利用できることによります。
チトクロムb/c複合体とその周辺の電子伝達系路はミトコンドリア・葉緑体ともよく似ています。細胞共生説から葉緑体の祖先種であると考えられるシアノバクテリアは、その内膜系上に光合成の電子伝達系路と呼吸の電子伝達系路の両方を持っています。両者の電子伝達系では、「プラストキノン-チトクロムb/f複合体(ミトコンドリアのb/c複合体に相当)-チトクロムc(高等植物のプラストシアニンに相当)」が共通になっています。
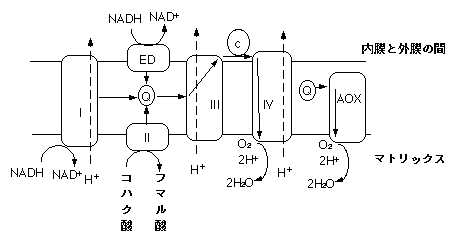
植物ミトコンドリアの電子伝達系の模式図。実線が電子の流れ、破線がプロトンの流れ。ATP合成酵素は省略。EDは外部NADHデヒドロゲナーゼ、AOXはオルタナティブオキシダーゼ、Qはユビキノン、cはチトクロムc。
植物のミトコンドリアには動物のミトコンドリアにはない経路がいくつかあります。植物は上記複合体I・II・III・IV以外にいくつかの複合体を持っており、これらが特殊な電子伝達経路に携わります(I・II・III・IVはウシのミトコンドリアから発見されたので、植物ミトコンドリアの複合体に通し番号?はない)。
・外部NADHデヒドロゲナーゼ(external dehydrogenase)
外部NADHデヒドロゲナーゼは、NADHデヒドロゲナーゼ(複合体I)同様NADHから電子を受け取り、ユビキノンに電子を渡します。複合体Iと違うところは、第一にNADHを分解する場所です。複合体Iはマトリックス側(内膜内側)で分解しますが、外部NADHデヒドロゲナーゼは内膜の外側で分解します。NADHは内膜を通過しないらしいので、外部NADHデヒドロゲナーゼが使うNADHはマトリックス由来ではなく、細胞質内に存在するNADHです。第二点は、プロトン輸送を行わないことです。
外部NADHデヒドロゲナーゼの存在意義は、細胞質内の酸化還元レベルの調節にあると考えられています。何らかの原因で細胞質が過剰に還元的になった場合、NADHを安全に(ATPを生産せずに)消去する機能があると考えられているようです。そうとうNADH濃度が高くならないと外部NADHデヒドロゲナーゼは機能しないようです。
外部NADPHデヒドロゲナーゼというのもあるらしいんですが、よくわからないので書かないでおきます。
・オルタナティブオキシダーゼ(alternative oxidase)
alternativeという単語は訳しにくいです。金井龍二先生は「代替オキシダーゼ」と忠実に日本語に訳していますが、イマイチ物質の名前に見えないところが難点です。それはともかく、オルタナティブオキシダーゼはシアン耐性呼吸の主要経路として知られています。まずはシアン耐性呼吸からお話ししましょう。
私がかなり小さかった頃、毒入りコーラがあちこちの自販機に入っている、という事件がありまして、そのとき使用された毒が青酸カリ(シアン化カリウム・KCN)でした。以来しばしば話題になるポピュラーな毒性物質です。KCNはチトクロムオキシダーゼ(複合体IV)の活性を阻害するので毒なわけです。KCNを動物細胞に与えると呼吸が止まります。しかし植物では止まりません。シアン存在下でも起こる呼吸ということでシアン耐性呼吸と呼ばれるようになりました。
シアン存在下でも呼吸が起こるのはチトクロムオキシダーゼと別にもう一つオキシダーゼがあるからです。それがオルタナティブオキシダーゼです。オルタナティブオキシダーゼはチトクロムcから電子を奪い、酸素を還元して水にします。ただし、チトクロムオキシダーゼと違ってプロトンの輸送はしません。
オルタナティブオキシダーゼやシアン耐性呼吸の意義は必ずしもはっきりしているわけではありませんが、これもやはり過剰な還元力の消去のためにあると考えられているようです。オルタナティブオキシダーゼはプロトン輸送をしませんので、無駄にATPを生産することなく還元力を消去できるわけです。
シアン耐性呼吸の生態学的意義が明らかな例は一つだけ知られています。サトイモ科の一部の植物では、花序においてシアン耐性呼吸の活性が上がります。オルタナティブオキシダーゼはATP合成に関与しないため、余ったエネルギーが熱になり、花序の温度を増加させます。これによって芳香物質が飛散しやすくなり、訪花昆虫を引き寄せるのに役立っているとされています。
なお、「シアン耐性経路が動くと全くATPを生産しなくなる」わけではないことに注意して下さい。。仮にチトクロムオキシダーゼの活性が0になり、全ての酸素消費がオルタナティブオキシダーゼで起こったとしても、チトクロムcに電子が渡るまでに複合体IやIIIでプロトン輸送が起こっています。
シアン耐性呼吸は植物だけにあるのではなく、一部の昆虫などにもあることが知られています。どういう意味があるのかは、私は知りません。エサとなる植物がシアン化合物を持っている場合、それに対抗するためなんでしょうか?