光合成の生理生態学講座
はじめに(200201 31)
葉フェノロジー(200201 31)
葉の寿命のバリエーション(200202 15)
コストベネフィット(200203 01・200203 16)
栄養塩利用(200204 10・200407 18)
葉の寿命は様々です。針葉樹では10年近く生存する葉もありますし、成長の速い草本では数週間で落ちてしまうものもあります。落葉樹のように、植物の「見た目」に大きな影響を与えることもあり、興味を惹きやすい話題です。ほっといても面白い話なんですが、葉の寿命が植物の重要な戦略であることが指摘されて以来、学術的にホットな問題となっています。
フェノロジー(phenology)という言葉は、あえて訳すと「植物季節」という言葉になるらしいですが、わざわざ訳して使っている人は少数です。季節に依存した植物の性質の変化をフェノロジーと呼んでいます。葉の着脱が季節にどう依存しているか、を調べた人が、「葉のフェノロジーを調べました」というように使います。
常緑種とは読んで字のごとく常に緑色、つまり葉を持っている植物です。落葉種はある季節に葉を落とす植物です。乾燥地には半落葉種というのもあるそうです。乾季に持っている葉の半分程度を落とすからそう呼ばれているのだと思いますが、よくは知りません。
落葉種が落葉するのは、その季節が光合成に不適であるからと考えてよいでしょう。乾燥地の乾季、温帯・寒帯の冬季がこれにあたります。一時期全ての葉を落とすわけですから、落葉種の葉の寿命は全て一年よりは短くなります。
常緑種の多くは長い葉の寿命を持ちます。しかし全ての種の葉の寿命が一年より長いわけではありません。熱帯では季節がないため、ほとんどの植物が常緑性です。葉の寿命が数ヶ月の植物でも、絶えず新しい葉が出れば常緑に見えるわけです。Reich et al. (1991) はアマゾンの熱帯林で23種の樹木の葉の寿命を測定し、数ヶ月から数年まで多様であったことを報告しています。
温帯でも、数は多くありませんが、常緑かつ一年未満の葉寿命を持つ植物はいます。私の知っている例では、アオイスミレがそれで、夏には夏葉(かよう)を冬には越冬葉を持ちます。それぞれの季節が終わるとそれぞれの葉は枯れます。フェノロジー屋さんは「入れ替え型常緑植物」と呼んでいます。
落葉種では、生育に好適な季節が来ると葉を展開します。このとき、多くの葉を一度に作ってしまい、その後は葉の数を増やさない植物と、最初に少数の葉をつけ、徐々に葉の数を増やしていく植物が見られます。前者を一斉展葉型、後者を順次展葉型と呼びます。もちろん中間型も存在します。一斉展葉型の植物は葉を落とすのもほとんど一斉で、葉の寿命は好適な期間の長さとほぼ等しくなります。順次展葉型もやはり葉を順次に落とします。したがって葉の寿命は好適な期間よりも短くなります。順次展葉型には成長の速いカバノキ科のような植物があり、一斉展葉にはブナなど成長が遅い種が多いようです。このようなタイプ分けは古くからあったようですが、メジャーな問題にしたのは菊沢喜八郎氏(現京大)の論文からではないかと思います(Kikuzawa 1983, 1984)。
このようなタイプわけは落葉樹に用いられることが多いのですが、ほとんどの高等植物はどちらかに分類することができます(葉を着けない植物はできませんが)。一年草など明るい場所を好む植物はほとんどの場合順次展葉ですが、林床草本などの中には一斉展葉的なふるまいを示す植物が多いようです。熱帯でも、展葉が周期的に起こる種と、連続的に起こる種がいるようです。
キソウテンガイ(正しい和名はサバクオモトとかいうらしいですが)という植物は、個体の寿命が数百年あるらしいのですが、この植物は生涯で葉を二枚しか作りません。しかし葉は数百年生き続けるわけではなく、絶えず根元から伸び続け、先っぽから枯れていく、というのを続けているのだそうです。
まあキソウテンガイは例外の部類に入ると思います。多くの植物では葉に寿命があります。寿命のバリエーションは種によって様々です。短いものだと、水面に葉を浮かべている植物の中には2週間程度の寿命を持つものがあるようです(Kikuzawa and Ackerly 1999)。逆に、長いものでは、数十年に達します。私が持っている文献の中では20年に達するものがあるようです(Reich et al. 1992)。
種間で比較すると、葉の寿命は様々な生理生態学的パラメータと相関があります。例えば、葉の寿命が長い種ほど、低い光合成能力・低い窒素濃度・高い葉重/葉面積比を持つ傾向があります。これについては「光合成能力の種間差」に詳しく書きましたので、ご覧下さい。この相関は様々な生態系の植物に共通して見られる傾向であることが示されています(Reich et al. 1997)。
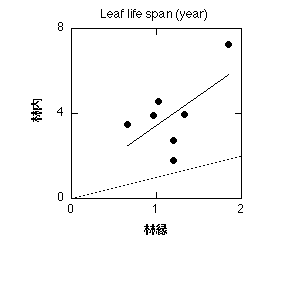
同一種でも、生育環境によって葉の寿命が変化します。よく知られているのは、貧栄養条件で生育した植物の葉の寿命が長くなるというものです(例えばAerts et al. 1995)。他にも、弱光環境で生育した植物で葉寿命が長くなるというのがよく知られています。ただ、「よく知られている」わりには、私はあまりそういう報告をしている論文を知りません。引用するときにはKikuzawa (1989) とかChabot and Hicks (1982) を使っているのですが、どちらもレビューで、著者が持っている予備的データが載っている、という程度です。探せばあるんでしょうけど・・・。私が持っている未発表データを載せておきましょう。東大千葉演習林で林縁と林内の常緑広葉樹7種の寿命を調べたものです。一点一種で、林縁では半年から2年なのに対し、林内では2年から8年と3~4倍長くなっています。
林内の異なる光環境に生育する葉の寿命。同一種について、林縁で生育している個体の測定データに対し林内で生育している個体の測定データをプロット。Hikosaka (2005) に同じデータを載せました。
種間比較をするときには種内変異に注意する必要があります。例えば、Reichらの一連の仕事(Reich et al. 1991, 1997など)できれいな相関が得られている理由の一つは、彼らが光条件が均一な葉を選んでいるからです。
弱光環境や貧栄養で葉の寿命が長い、というのは、葉の老化の知見と必ずしも一致しないように見えます。葉の老化は光環境の悪化や栄養塩不足で促進されると考えられているからです。この矛盾は、光環境については私の想像をこちらに書きましたのでご覧下さい。栄養条件の問題についても、いずれ書きたいと思います。
葉の寿命については不思議なことが多いです。例えば、なぜ寿命の長い種と短い種がいるのか。また、なぜ常緑種と落葉種がいるのか。それぞれにそれなりの理屈をつけることはできます。しかし、これらの種が同一環境に共存できるとなると、理解は難しくなります。例えば、片方の形質が有利であることを示すことは、暗に別の形質が不利であることを意味します。ならば、一つの環境にはどちらかの形質しか存在しないはずです。また、わかりやすい謎として、常緑と落葉種の緯度別の分布があります。低緯度ほど常緑種が多くなりますが、中緯度域では落葉種が多くなり、高緯度ではまた常緑種が多くなります。こういった複雑な分布を一つの理論で説明できるでしょうか? ここではこれらの問題についての先人達の努力を紹介します。
「コストベネフィット」そのものは生物のふるまいを、そのコスト(必要なエネルギーや投資資源)と利益(獲得できる資源や残せる子孫の数)を定量化することで理解しよう、というものです。生理生態のレベルでは、古くはBoysen-Jensen (1932) に始まり、Mooney and Gulmon (1979) から世界的にメジャーな概念となりました。
葉の寿命の問題にコストベネフィットの概念を持ち込んだのは、当のMooney and Gulmon (1982) とChabot and Hicks (1982) だと私は考えています(どちらかというとはっきり言っているのはChabot and Hicks のほうです)。彼らの考えを簡単にまとめると、「長い葉の寿命は低い光合成速度を補償するため」ということです。当時から、特に種間では光合成能力が低い種ほど葉の寿命が長い傾向があることがわかっていました。どうして光合成能力が低い種がいるのかはとりあえず置いておいて、そういう種の葉は自分自身(新しい葉)を作るためにある程度の炭素を獲得しなければならず、長い寿命をもつはずだ、と考えたわけです。実際に、「葉が一生に稼ぐ光合成量」を計算してみると、光合成能力が低い葉も高い葉もそれほどかわりがないことが指摘されています(Chabot and Hicks 1982)。とはいえ、この考え方では「なぜ光合成速度が高い葉では寿命が短いのか」は答えられないので、全てを説明できるわけではありません。まあ、「経験的な解釈」にとどまっていたわけです。ただし、Mooney and Gulmon 1982は、一年草では新しい葉が次々と生まれ、古い葉が被陰されていき光合成できなくなるために葉寿命が短いのだ、という解釈と組み合わせることによって説明しています。どちらにせよ、一つの理論で全てを説明できるわけではありませんでした。
葉の寿命の長短を一つの理論できれいに説明してみせたのがKikuzawa (1991) です。ここには一つ発想の転換があります。以前の考え方では、葉の一枚の炭素収支ばかりが問題になっていましたが、Kikuzawa (1991) では、「個体の炭素収支」を最大にする葉の寿命が検討されました。Kikuzawaモデルでの仮定は以下の通りです。1)葉の光合成速度は展葉直後に最大値aを持ち、葉寿命とともに直線的に減少する(ある時間bが経つと0となる。2)個体が持つ葉の数は一定である(ある数の葉が生産されると同数の葉を落とす)。3)新しい葉を作るためには一定のコストCがかかる。以上のような仮定から一枚の葉の炭素収支を考えると、以下の図黒線になります。
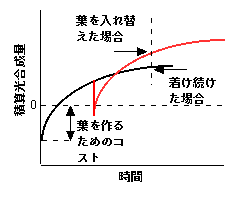
Kikuzawa (1991) モデルにおける葉の炭素収支のイメージ。もともとの論文にほとんど同じ図が載っています。ここで出したのは講義で使用したものに手を加えたものです。
縦軸は、葉の瞬間光合成値ではなく、積算値です。最初に葉を作るのにコストが必要なため、その分炭素収支はマイナスからスタートします。その後葉は光合成をするために炭素収支はプラスになります。しかし、時間が経つにつれて光合成速度が下がりますので炭素収入の増加が小さくなり、やがて増加が0になります。これに対し、ある時間で葉を入れ替えてみましょう(赤線)。新しい葉を作ると、直後はやはり炭素収支が大きく下がりますが、新しい葉のほうが光合成速度が高いため、やがて収支は葉をつけかえない場合よりも高くなります(図中矢印)。つまり、葉を入れ替えた方が得なわけです。
個体の生産を最大にする最適な葉の寿命t*は、計算して解くと以下の解となります。
t* = (2bC/a)1/2
解は個体が持つ葉の量には依存しません。最適な葉の寿命は、コストが高く、光合成が0になるような葉寿命が長くなるほど長く、最初の光合成速度が高いほど短くなります。これらの傾向は、野外でとられた経験的な傾向と一致します。
この数式の意味を直観的に説明すると以下の通りになります。まず、葉を作るコストが多い葉では、ある程度葉の寿命を長くしないと、次の葉を作るための材料を稼ぐことができません。逆に、光合成速度が高い葉では、次の葉の材料を比較的短期間で稼げるので葉の寿命が短くなります。光合成が0になるような葉寿命が長い、というのは、光合成速度がなかなか落ちないことを意味します。光合成速度が落ちないのなら、わざわざコストをかけて新しい葉を作る必要はありません。ので、葉の寿命が長くなります。このように書くとわかると思いますが、このモデルでは「個体が持つ葉の量が決まっている」ということが重要な仮定になります。個体が葉を無限に持つことができるのなら、全ての葉の寿命を最大限にするほうが個体の生産は多くなるからです。では個体が持つ葉の量がどう決まっているのか、についてはこのモデルでは考えていません。この点がKikuzawaモデルの次のステップであるのではないかと個人的には考えています。
Kikuzawa (1991) では、さらにこのモデルを使って落葉種と常緑種の緯度分布の説明を試みました。上に書きましたように、ある地域における常緑種の割合は、低緯度で多く、中緯度で少なく、高緯度で再び多くなるという二山型の分布を示します。この傾向を説明するために、菊沢さんは非常にユニークな考え方を導入しました。a、b、Cの三つのパラメータの数字の組み合わせを種の特性と考えたのです。例えば、それぞれのパラメータは1から10までの整数値をとると仮定しましょう。そして、a=1, b=1, C=1を種1とし、a=2, b=1, C=1を種2、・・・a=10, b=10, C=10を種1000というように、パラメータの組み合わせの違いで種を定義しました(ここで示したのはあくまで例です)。Kikuzawa (1991)では、冬期間の光合成速度が0であると仮定し、それぞれの「種」の最適な寿命を計算しました。そうすると、低緯度(冬期間が短い)では常緑種が多く、中緯度では落葉種が多くなり、高緯度では常緑種がちょっと多くなる、という二山分布が再現できることを示しています。ついでに書くと、Kikuzawa (1995) では上のように定義された「種」の数の緯度分布について検討しました。その環境で光合成生産をプラスにできる「種」は生き残ることができるが、できない「種」生きていけない、と考え、ある環境で光合成生産をプラスにできるパラメータの組み合わせの数を数えたのです。この結果、低緯度ほど光合成生産をプラスにできる組み合わせ数が多いことを示し、「なぜ熱帯は種数が多いか」を説明できる仮説としています。「面白い」説であるとは思います。
このモデルで予測される最適な葉寿命が現実の葉の寿命とどれだけ合うのか、という厳密な意味での定量的な比較は、私が知る限りではなされていません。しかし、Kikuzawa and Ackerly (1999) では、若い葉の飽和光で測った光合成速度をaとし、葉齢と光合成速度の関係を直線回帰してbを求め、その結果計算した(2bC/a)1/2と実際の葉の寿命の間に高い相関があることを示しています。また、Kikuzawa and Kudo (1995) では、雪田植生の植物の葉の寿命をこのモデルによって説明するという試みをしています。雪田とは、高山・亜高山地域などで、地形と気象の関係により積雪が他の場所よりも長く続く場所を指します。ここでは雪解けが遅いため、植物の生育期間が短くなります。ただし、生育期間中の環境条件には他の場所と大きな違いがないため、環境条件の影響を排し、植物に対する「生育期間の長さ」の違いの影響のみを評価できることがユニークな実験対象です。Kudo (1992) では、生育期間の長さが異なる地点に生育する落葉と常緑の低木の葉の寿命を報告しています。落葉種では、生育可能期間が長いほど葉の寿命が長いのに対し、常緑種では生育可能期間が長いほど葉の寿命が短くなります。この傾向はKikuzawaモデルの予測とよく一致しました。このあたりの背景は私の駄文より、工藤さんのご著書(工藤 2000)に興味深く書かれていますのでそちらをどうぞ。
上で紹介した研究は炭素収支にのみ着目した研究ですが、寿命と栄養塩利用に着目した研究も少なくありません。葉の寿命が長いということは、光合成を長く続けられるということと別の利点があります。葉が枯れる際には、ある程度栄養塩が回収されるとはいえ、数10%は回収されることなく枯葉とともに植物体から失われていきます。つまり、葉の寿命が長い植物ほど枯葉を捨てる機会が少なく、栄養塩を失いにくいというわけです。
このアイディアを初めて示したのはSmall (1972) です。Smallは、一枚の葉レベルでの窒素の利用効率を考えました。ここでは、葉に新たに投資された窒素(回収される分は含まない)あたりの光合成生産を考えました。このパラメータを高めるためには、光合成速度を高めるか、投資された窒素を長く保持するか、回収効率を高める、のいずれかをする必要があります。彼はこのパラメータを常緑・落葉種合わせて28種で比較しました。すると、瞬間的な光合成速度が高い落葉樹よりも、葉の寿命が長い常緑樹のほうが窒素利用効率が高い傾向がありました。このことから、特に栄養塩が不足しがちな土地では、栄養塩利用効率が高めるために葉の寿命が長い種が有利ではないかと考えられます。この考え方はBerendse and Aerts (1987) の(個体レベルの)窒素利用効率の考え方に引き継がれます。窒素利用効率についてはまた別に紹介したいと思います。
近年は栄養塩利用効率から葉の生き死にを考える研究も出てきました。まずはFranklin & Agren (2002) から。この研究では、直接葉の寿命を考えているわけではありませんが、どのような状況で葉を枯らすと最適か、という問題を考えています。彼らが用いたのは群落光合成における最適窒素分配モデルと最適葉面積指数モデルです。最適窒素分配については詳しくは別に説明する予定ですが、特に重要なのは後者の最適窒素分配を考慮に入れた最適葉面積指数モデルです。これらを簡単に説明すると以下のようになります。 1)ある葉面積指数・ある葉窒素含量(土地面積あたり)を仮定すると、群落光合成速度を最大にする最適な窒素分配がある (Anten et al. 1995a)。最適窒素分配では、上部の葉ほど高い窒素含量を持つ。これは、単位窒素投資に対する光合成量の増加が、強光下の葉ほど高いためである(かみ 砕いて言うと、強光にある葉に多く窒素を投資すると投資した分だけ光合成速度が上がるが、弱光にある葉に窒素を投資しても光合成速度は光に律速されて上が らない。ならば上の葉に投資した方が有利)。2)土地面積あたりの葉窒素含量を一定と仮定し、窒素分配が最適に調節されていることを仮定すると、群落光合 成速度を最大にする葉面積指数が存在する (Anten et al. 1995b)。土地面積あたりの窒素は一定なので、葉面積を増やしすぎると、葉面積あたりの窒素含量が低くなってしまう。逆に、葉窒素含量を上げすぎると 葉面積指数が低くなってしまう。Anten et al. (1995b) のモデルでは、群落は葉面積を自由に変えられる、というのが仮定です。つまり、土地面積あたりの窒素量を一定にし、いろいろな葉面積指数で群落光合成速度 を計算し、群落光合成が最大になるような葉面積指数を探していたわけです。ここでFranklin & Agren (2002) は、葉面積指数の変化に一つ生物学的な制約を加えました。それは、「葉面積指数が減るときには窒素の減少を伴う」という制約です。ここでイメージしている のは、葉が枯れるときに、一部は回収されるが、残りは枯葉とともに植物体から失われる(=土地面積あたりの窒素量が減る)ということです。最適値よりも葉 面積指数が多い場合、葉面積指数を減らすわけですが、そのときには窒素量の減少を伴っているので、窒素回収を仮定するか仮定しないかで最適な葉面積指数が 変わってしまいます。Franklin & Agren (2002) は特に最下層の葉に着目し、「葉を枯らすことが群落光合成速度の増加をもたらすか否か」を計算し、最適な葉面積指数を計算しました。窒素回収効率(枯死ま でに回収された窒素量/葉の最大の保有窒素量)が高いほど最適葉面積指数が小さくなることを示しています。
この考え方を実際の植物に応用したのがEscudero & Mediavilla (2003) です。彼らはFranklin & Agren (2002) の考え方を単純化し、以下のような仮説を提案しました:「PNUE(窒素あたりの光合成速度)の減少率(その葉のPNUE/若い葉のPNUE)が窒素回収 効率を下回ったら葉は枯死する」。これは説明がまたたいへんなのですが、ざっと以下の通りです。1)PNUEは葉の寿命が進むとともに低下する(仮に、若 い葉のPNUEを100としましょう)。今、ある葉のPNUEが50まで低下してしまった。2)PNUEが低い葉を保持することは植物にとって得ではない かもしれない。では、古い葉から窒素を回収し、若い葉をつくればいいではないか? 3)しかし、全ての窒素を回収できるわけではない。仮に窒素回収効率が 30%だったとしよう。この葉を枯らすと30%の窒素を回収し、新たな葉を作ることができる。この葉では、送られた窒素はPNUE×窒素量=100× 0.3=30の光合成量を実現できる。4)ということは、いくら低下したとしてもPNUEが50の葉のほうがまだ高い光合成速度を維持できる。つまり、 PNUEが30に下がるまでは、この葉をつけておいたほうが良い・・・・。てな感じです。わかっていただけたでしょうか。要するに、枯れた葉をそのままつ けておいたほうが光合成量が多いのか、回収できなかった窒素を失ったとしても若い葉をつけたほうが良いのか、という選択なわけです。Escudero & Mediavilla (2003) は地中海性気候の低木数種のPNUEの低下と窒素回収効率を比較し、この考えが当てはまる種もあれば当てはまらない種もあることを示しています。
Franklin & Agren (2002) の研究は、最適葉面積指数に着目しているせいもありあまり時間変化に気を配っていません。これに対し最近発表した私のモデルは、比較的似たような理屈を使っているのですが(注)、時間変化を強く意識したものになっています(Hikosaka 2003)。モデルの仕組みはFranklin & Agren (2002) とそう大きく変わるわけではありませんが、こんな感じで計算しています(数学的にはダサいです)。
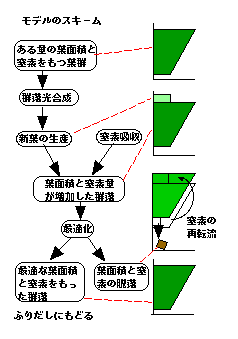
ある量の葉面積と窒素量をもつ葉群を考えます。この葉群は一定期間光合成をし、その炭素を使って新葉を生産します。同時に根から窒素を吸収し、新葉 に送ります。この時点で、葉面積と窒素量が増えているわけです。ここに最適葉面積指数モデルを適用します。もちろん、葉面積指数が減った場合には、枯葉と ともに窒素量が減ることが考慮に入っています。この結果葉面積と窒素が減った葉群ができます。これはまた光合成をし、窒素を吸収して・・・とループを繰り 返すことで葉群の成長をシミュレートするわけです。
このモデルでは群落光合成モデルを使っていますので、光環境の違いをモデルに組み込むことができます。また、新葉に送られる窒素供給速度を栄養条 件の反映とみなすことができます。また、窒素と光合成の関係を変えることでその他の環境の影響や種によるPNUEの違い(詳しくはこちら)をモデルに組み込むことができます。さらに葉の寿命や窒素利用効率をモデルの出力とすることができます。私としてはこのモデルは葉群の動態とその環境応答を包括したものだと自負しています(誰も引用してくれませんが)。
この研究ではいろいろなことがわかったのですが、葉の寿命に関するところだけ書きます。葉の寿命は、強光環境ほど短くなります。あと、LMA(葉 面積あたりの葉重量)が大きいと寿命が長くなります。これらはほとんどKikuzawaモデルと同じ理屈で説明できます。Kikuzawaモデルが適用で きないのは窒素にかかわることですが、私のモデルでは栄養条件が悪くなると寿命はちょっとだけ長くなります。ただし窒素の影響は複雑で、例えば長期間富栄 養で植物を育て、いきなり貧栄養条件にすると葉の寿命は一時的に短くなり、徐々に長くなったり、と実際の植物でも起こっていそうな変化も示します。
注:私のモデルとFranklin & Agren (2002) のモデルの一番違うところは、「葉を枯らすときのルール」です。Franklin & Agren (2002) のモデルでは、ある葉面積指数・ある窒素量を与え、最適窒素分配を計算し、その後で最下層の葉を枯らすか否かを考えているようです。これに対し、 Hikosaka (2003) では、時間t-1の状態としてある葉面積指数・ある窒素量を与え、ここから時間tいおける最適窒素分配を維持しつつ葉面積指数を変化させることによって最 適葉面積指数を計算しています。つまり最適葉面積指数を計算するステップに違いがあります。これが重要な違いなのかどうかは私にはわかりません(笑)。な お、私のモデルはFranklin & Agren (2002) とは独立に作ったものです。論文を投稿したときには彼らのモデルはまだ世に出ていませんでしたから。