光合成の生理生態学講座
はじめに(200410 19)
植物はどれだけ食べられているのか(200410 19)
植食者の餌としての植物(200410 31)
二次代謝物質(200411 22)
テルペン類(200411 22)
フェノール類(200411 28)
含窒素化合物(200412 16)
なぜ多様な防御物質が進化してきたか(200501 18)
量的防御と質的防御(200506 26)
炭素-栄養塩バランス仮説(200501 30)
成長-分化バランス仮説(200502 22)
資源量仮説(200504 03)
みてくれ仮説(200506 16)
「世界はなぜ緑なのか?」という有名な問いがあります。植物の葉は何らかの草食動物によって食べられることが頻繁にあります。ほとんどの場合、植物は逃げることはできませんので、放っておくと全ての葉を食べられてしまいかねません。実際、ある大面積の植生の葉が全て食べられてしまうような大発生もしばしば起きます。しかし、そのようなことはまれで、全体的に見ると陸域の多くの場所は植物で満たされており、「世界は緑」なわけです。「どうして植物は食べ尽くされずに存在することができるのだろう?」というのが植物-動物関係の基本的な疑問の一つです。というわけで、この項では植物の葉をめぐる「食う-食われる関係」にまつわる話を書きます。主としてHartley and Jones (1995) の内容を参考にします。
まずは基本的な問題として、生態系で植物はどれだけ食べられているのか、というのがあります。植物によって生産される生物量を乾燥重量として表したものは純一次生産と呼ばれます。このうち何割が草食動物によって失われたのか、ということを調べた研究例をまとめた研究があります(Cyr and Pace 1993)。面白いのは、生態系によってその比率が大きく異なることです。水界の浮遊藻類は、研究によってばらつきは大きいのですが、平均すると植物の純一次生産の80%が植物を食べる動物(植食者)によって食べられています(グラフを見る限り、純一次生産より植食者の生産が大きい例-系外の有機物由来?-も多いので注意が必要ですが)。これに対し陸域生態系では、多くの場合植食者が食べた量は20%より低く、平均値も18%にとどまります(こういった研究では被食量が多い生態系が選ばれている可能性は高いと思うので、実際の平均値はもっと低いことでしょう)。つまり、上に書いた「世界はなぜ緑なのか?」という問いへは、「あまり食べられていないから」というのが答えの一つとなるのだと思います。逆に、海や湖が緑でない理由は「食べられるから」、ということになります。
さて、ではなぜ陸上植物は「あまり食べられていない」のでしょうか? その理由は、動物にとって植物が餌として優れていないから、という点につきます。主に二つの要因があります。
一つは、植物の窒素濃度が著しく低いことです。植物の器官のうち、一般に最も窒素濃度が高いのは葉ですが、乾重あたりで5%を超えることはめったになく、常緑木本では1%を下ることも珍しくはありません。茎・根といった器官の窒素濃度は葉の窒素濃度の半分以下になるのが普通です。これに対し、動物の窒素濃度は多くの場合10%を超え、おおざっぱに言って植物の窒素濃度の10倍です。この違いは主として植物が細胞壁の繊維などの炭素化合物を多量に持っていることによります。つまり動物の餌としては炭素が過剰で窒素が不足しているということです。植食者の成長は基本的に窒素の摂取量に律速されることになります。仮に植食者が食べた餌の窒素を100%吸収・利用できたとしても、植食者が1g成長するためには10gの葉を食べなければならないことになります。これはいかに植物が動かないとはいえ、非常に効率が悪いと思えます。逆に、植物の窒素濃度が上がれば、植食者にとっては有利です。植物に施肥をすると、それを食べた植食者には成長増加などのポジティブな効果が出ることが多いようです(Waring and Cobb 1992)。
このような議論は窒素だけでなくリンなど他の元素にも当てはまると考えられていますが、あまりよく調べられていないようです。
もう一つは、植物が植食者に食べられにくいように工夫をしていることです。トゲや毛を作る物理的な防御と「まずい」物質を体内にため込む化学的な防御に大別することができます。化学的防御としてため込まれる物質は二次代謝物質(secondary motabolite, secondary compound)などと総称されます(実際には、「二次代謝物質」の全てが化学防御にかかわっているわけではありませんが、防御にかかわっている二次代謝物質が非常に多いので、「二次代謝物質」=「化学防御物質」と扱われることが多いです)。現在数万種類?の防御物質が知られているようです。それぞれの防御物質は、毒、消化を妨げる、忌避など多種多様な機能が知られています。これらの物質の蓄積により植物は食べられにくくなっています。
では二次代謝物質の生理学について簡単に説明しましょう。私もほとんど知識がないので、Teiz and Zeiger (2002) の要約程度にしかならないと思いますが。
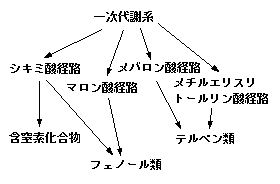
二次代謝物質は、大きく分けて三つのグループに分かれます。テルペン類、フェノール類、窒素化合物の三種類です。
テルペン類(terpenes)あるいはテルペノイド、イソプレノイドなどと呼ばれます。テルペン類にはジベレリンやアブシジン酸といった植物ホルモンやカロテノイドといった色素もこのグループに含まれますが、こういった物質は成長に必須なので、一般には二次代謝産物とは言いません。我々が普段の生活で耳にするようなテルペン類は、例えばサポニンとか、精油(芳香油とも。ペパーミント、レモン、バジルなどの香りの物質)、ジギタリス(正確には、ジギタリスという植物に含まれるカルデノライド)が挙げられます。
全てのテルペン類は以下のようなイソペンタンを一つの単位とし、これが重合することによって合成されます。
H3C-CH-CH2-CH3
|
CH3
この単位をイソプレン単位と呼ぶそうです。これはテルペン類を高温によって分解するとイソプレンが生じるためだそうです。2つのイソプレン単位からなるテルペン類(つまりC10)をモノテルペン類、3つのイソプレン単位からなるテルペン類を背好きテルペン類、4つだとジテルペン類、6つだとトリテルペン類、8つだとテトラテルペン類、それ以上だとポリテルペン類と呼ばれます。
テルペン類の合成経路は二種類知られています。よく知られているのが、メバロン酸経路です。TCA回路の中間代謝産物であるアセチルCoAが3分子結合してメバロン酸が合成され、イソペンテニル二リン酸(IPP、C5)が合成されます。ここまでがメバロン酸経路です。もう一つの経路はグリセルアルデヒドリン酸やピルビン酸からIPPが合成される経路で、メチルエリスリトールリン酸経路と呼ばれます。IPPがさらに重合することにより、高分子のテルペン類が合成されます。
植食者に対するテルペン類の作用は物質によって様々で、体系的に説明するのは難しそうです。毒性があるもの(ピレスロイド、カルデノライド)、忌避物質(精油、リモノイド)、ホルモン様物質(ファイトエクジソン)などいろいろです。面白いものでは、植食者に食べられたあとに合成される揮発性テルペン類があります。これは植食者に直接作用するのではなく、植食者の天敵や寄生虫を誘因して植食者を撃退するのだそうです。
フェノール類(phenolics)、あるいはフェノール性化合物などと呼ばれます。この物質はフェノール環(ベンゼン環の外側の水素の一つが水酸基に置換したもの)が基本構造です。
植物では、フェノール類物質のほとんどはシキミ酸経路から生合成されます。カビやバクテリアではマロン酸経路が主流なのですが、植物ではほとんど使われていないようです。シキミ酸経路はペントースリン酸経路の中間代謝産物エリトロース-4-リン酸か解糖系の中間代謝産物ホスホエノールピルビン酸(PEP)から出発し、最終産物はフェニルアラニン(ベンゼン環を持つアミノ酸)です。フェニルアラニンからアミノ基が取り除かれ、さらなる生合成を経て様々なフェノール類物質が合成されます。なお、ほとんどのフェノール類物質は窒素をもたないようです。
フェノール類物質は10000種類ほどが知られているようです。どう分類するのかは本によって違うようですが、Teiz and Zeiger (2002) にしたがうと、1)単純フェノール化合物、2)リグニン、3)フラボノイド、4)タンニンの4種類に分類できるようです。
単純フェノール化合物はフェノール環を少数持つ(比較的)低分子化合物です。クマリンや安息香酸などが知られています。これらの物質は害虫やカビへの防御化合物としてはたらいているようです。
リグニンはフェニルプロパノイドというフェノール環に炭素が三つついている物質が高度に重合してできた高分子です。細胞壁中のセルロースなどと共有結合し、細胞壁を物理的に丈夫にする役割を持っています。植物が含有している物質としてはセルロースの次に多い物質です。
フラボノイドはフェノール類で最も多様です。フラボノイドはさらに分類することができ、アントシアニン、フラボン、フラボノール、イソフラボンの4種類に分けられます。植物の色素の多くはテルペン類の一種であるカロテノイドとこのフラボノイドに大別できますが、有色のフラボノイドのうち最も広く存在するのがアントシアンです。アントシアン類は赤・ピンク・紫・青など様々な色を持っており、訪花動物の誘因に役立っています。ただ、防御に有効かどうかは知りません。フラボンとフラボノールは紫外線を吸収します。ハチのような昆虫は紫外領域の光を感じるため、これらの物質も誘因に役立っています。また、フラボン・フラボノールは葉・茎の表皮細胞に蓄積されており、葉の内部を紫外線から守るはたらきを持っています。ただし、これらも防御に有効かどうかは知りません。イソフラボノイドはマメ科植物に多く含まれ、強い抗菌活性を持っています。
タンニンは、縮合型タンニンと加水分解型タンニンの二つに分類されます。縮合型タンニンはフラボノイドに由来する高分子物質です(フラボノイドを構成する単位が重合することによってできている)。加水分解型タンニンは、安息香酸類の一種である没食子酸と単糖が重合してできています。タンニン類は、植物が食べられるとタンパク質と結合して複合体を形成し、消化されないようにすることで草食動物の成長を阻害します。また、「渋味」があり、忌避効果もあります。
テルペン類もフェノール類も基本的にはCHOの3元素で構成される物質ですが、窒素を含む二次代謝化合物もあります。ここではこれらを含窒素化合物として一くくりに扱います。含窒素化合物は、アルカロイド、シアン配糖体、カラシ油配糖体、その他に大別できます。
アルカロイド
アルカロイド(alkaloids)はアミノ酸から合成され、複素環(ヘテロ環)と呼ばれる環状構造を持ち、窒素原子はこの環状構造の中に含まれます。アルカロイドは全ての植物に含まれるわけではなく、維管束植物全体の2割に見出され、15000以上の種類が知られています。よく知られているものにはモルヒネ、ニコチン、カフェイン、ストリキニーネ、スコポラミンなど日常生活で耳にすることもあります。アルカロイドの多くは神経伝達系に作用し、少量で強い毒性があります。
シアン配糖体
シアン配糖体(cyanogenic gulycosides)はシアン(-CN)と糖の化合物で、それ自体は必ずしも毒性ではありません。シアン配糖体からグリコシダーゼによって糖が切り離され、さらにヒドロキシニトリルリアーゼという酵素によってヒドロキシニトリルもしくはシアノヒドリンという加水分解物が生じ、これがゆっくりと分解されることによってシアン化水素(HCN)が遊離してきます。HCNは呼吸阻害などを引き起こします。
シアン配糖体はマメ科やバラ科など様々な植物で見られます。これらの植物はシアン配糖体と分解酵素を別の部分に持つことによって通常はシアンの生成が起こらないようにしています。ソルガムという植物では、シアン配糖体が表皮細胞の液胞に、分解酵素が葉肉細胞に局在しており、通常はシアン配糖体の分解は起こりません。補食によって葉が傷つくと、両者が混じり合って分解が進むことになります。
カラシ油配糖体
カラシ油配糖体(mustard oil glycosides)は, グルコシノレート(glucosinolates)とも呼ばれ、アブラナ科とその近縁な植物種に見られます。これもやはり配糖体から糖が外れ、イソチオシアネートやニトリルといった毒性のある物質が生じます。
その他
・非タンパク質アミノ酸・・・捕食者体内でアミノ酸合成や輸送を阻害
・レクチン・・・タンパク質の一種。炭水化物などに結合して消化を阻害する。
・タンパク質分解酵素阻害剤・・・消化を阻害。
上で示したように防御物質の種類はおそらく数万種に達します。なぜこのように多様なのでしょうか? 説をいくつか紹介します。
1 共進化(coevolution)
まず、植物の適応度が被食に影響を受けている、ということが前提です。しかし植物は突然変異などによって新たな防御物質を生産できるようになることが(まれに)あります。そうなるとその植物は被食されなくなり、子孫を多く残すことができるようになります。一方、植食者も分解系の獲得などによりその防御の効果を受けないような進化をすることがあります。そうなると新たな防御物質を生産できるようになった植物が食われるようになります。そこで、また新たな別の防御物質を生産するような植物が、というように、植物と植食者が新たな防御と解毒の方法を進化させているために防御物質は多様なのだ、という考え方です(Ehrlich and Raven 1964)。このような場合、しばしば植物の系統樹とそれを食べる植食者の系統樹はよく似た形になります。異なる集団の個体の性質が別の集団の個体の性質との相互作用によって起こる進化を共進化と呼びます。被食系だけではなく、訪花系など様々な系で見られます。
2 結果として起こる進化(sequential evolution)
※適当な訳を知りません(ご意見お待ちしてます)。
共進化に近いですが、植物の防御物質の多様さは植食者の影響を受けたためではなく、別の理由(例えば環境応答とか植物-植物間相互作用とか)だとする考え方です。一方、防御物質の性質は植食者の進化には影響を与えると考えています(Jermy 1976)。
3 スクリーニング仮説(Screening hypothesis of chemical diversity)
新たな防御物質の生合成経路をもたらす突然変異というのはそう簡単に起こるわけではありません。実際には、既存の生合成経路に何らかの変異が起こって前とはちょっと違う物質が作られるようになる、という程度のものでしょう。したがって新たな防御物質の生産の進化を容易にするためには、(その結果生産される物質が現在必ずしも防御に有効とは限らなくても)様々な生合成経路を保有しておかなければいけません。そういう可能性を残していた植物が現在まで生き残っていて、その結果として多様な防御物質が存在している、ということです(Jones and Firm 1991)。
タンニンなど防御物質の濃度は一定ではなく、同一種でも生育条件などによって大きく変動します。どのようなときに濃度が高く/低くなるのか、この濃度変化をどのように説明するのかについていくつか理論があり、一部ではアツい議論があります。いくつか紹介します。
これは理論というほどのものではありません。上に述べたように二次代謝物質は多様で、植食者に対する効果のメカニズムもまた多様です。タンニンなどの物質はタンパク質にべたべたくっついて消化を妨げるという効果があります。このような物質は少量ため込んでも効果は少なく、量が多ければ多いほど効果が大きくなります。このような物質を蓄積することを量的防御と呼びます。一方、あるものはホルモン様の物質で植食者の内分泌を撹乱し、あるものは基礎代謝を阻害する「毒」の作用を持ちます。こういった物質は比較的少量で効果があるため、量を多くためこむ必要がありません。このような防御を質的防御と呼びます。質的防御のメリットは防御物質の生産にコストがあまりかからなくてすむことですが、植食者側が解毒能力を持つと防御の効果がなくなってしまうというデメリットがあります。
炭素-栄養塩バランス仮説(carbon-nutrient balance hypothesis)
Bryant et al. (1983) によって提示された仮説です。CNB仮説と略され、その理解の容易さもあり80年代から90年代にかけて被食応答の研究で非常に着目された仮説です。簡単に説明すると以下のようになります。植物の成長はほとんどの場合炭素もしくは栄養塩の供給に律速されています。片方、例えば栄養塩が不足していれば、植物体内では炭素が過剰になります。このような場合、余った炭素からタンニンなどCHOのみで構成される(Nを含まない)防御物質を合成すれば過剰な炭素を効率よく利用できることになります。一方、逆に窒素が余った場合はアルカロイドなどの含窒素化合物を合成すれば効率的と思えます。
この仮説は施肥(栄養塩供給を増やす)や被陰(炭素供給を減らす)などによって検証することが可能で、実際様々な研究者によって検証が行われ、例えば、Bryant et al., 1987a, b; Price et al., 1989; Gebauer et al., 1998; Hemming and Londroth, 1999; Dormann, 2003などが施肥によってフェノール類の濃度が減少することを確認しています。しかしその一方で予測に合わない結果も多く、妥当性に疑問がもたれるようになりました。決定的だったのはKoricheva et al. (1998) によるメタ解析(多くの文献の結果を集めて行う統計解析)による報告です。彼らは炭素ベース化合物(Nを含まない)をフェニルプロパノイド(加水分解性タンニンを除くフェノール類)、加水分解性タンニン(このページの分類では加水分解性タンニンもフェノール類に入れていますが、合成経路が他のフェノール類とちょっと違うのでこの解析では別に分類されています。なお、この解析では縮合性タンニンはフェニルプロパノイドに含まれていると思います)、テルペン類の3種類に分け、これらの化合物濃度に対する植物の生育環境6種類(窒素・リン・光・CO2・乾燥・オゾン)の変化の影響を解析しました。その結果、窒素施肥、被陰、CO2処理は炭素ベース化合物の濃度を仮説の予測のように有意に影響していることが示されましたが、3種類に分けて解析してみると、有意に影響されているのはフェニルプロパノイドだけで、加水分解性タンニンやテルペン類は影響されていませんでした。Koricheva et al. (1988) はCNB仮説は防御に使える炭素の量を説明できるが、個々の物質の量までは説明できないのだ、としています。また、出典は忘れましたが、窒素ベース化合物の濃度はCNB仮説ではほとんど説明できないと聞いたことがあります。2001年には「CNB仮説を棄却すべきだ」とする論文も出ました(Hamilton et al. 2001)。2002年にはOikos誌(これはBryant et al. 1983を掲載した雑誌)にCNB仮説をどう扱うべきかの論争が掲載されています(Lerdau and Coley 2002, Koricheva, 2002; Nitao et al., 2002)。
私の個人的見解を述べておきます(専門家ではないので、英語で主張する気にはちょっとなりませんが)。私はフェニルプロパノイドしかCNB仮説に合う化合物がないとしてもCNB仮説を捨てる必要はないと考えます。まず、フェニルプロパノイドだけでもかなりの種類の化合物、特に重要な縮合性タンニンを含みます。もう一つは、炭素-栄養塩バランスが重要なのは防御物質濃度の話だけではないことです。例えば地上部/地下部比のように、炭素と窒素のバランスによって変化する生理・生態的特性は数多くあります。これらの応答と防御物質濃度応答をリンクさせて考えるときにCNB仮説は重要な意味を持つと思います。地上部/地下部比も、窒素施肥に対しては応答するけどCO2濃度上昇に対しては応答しないなど、単純に炭素と窒素のバランスだけで説明できるわけではありません。しかしこのような考え方がその後の解析のベースとなることは有用だと思います。
成長-分化バランス仮説(Growth-differentiation hypothesis)
GDB仮説と略されます。植物が獲得した資源の使いみちを細胞レベルで考えると、その資源を(1)細胞の分裂や伸長に使うか(2)細胞の成熟や特化に使うか、という選択肢があります。簡単に言って、大きくなるため(成長)に使うか、防御に使うか、という二者択一です。隣接個体との光競争に勝つためには成長が重要だし、食べられないためには防御が重要です。状況によってどちらにより多く投資すれば良いかが異なり、その結果防御への資源投資比率が異なる、というわけです(Herms and Mattson 1992)。
この説は、もともとCNB仮説が防御物質の挙動を説明できないケースが多いため代替仮説として提案されました。GDB仮説はCNB仮説を包含し、CNBでカバーできない領域も含むのでCNBの欠点を補うものとして提案されました。ただ、CNB仮説と矛盾するわけではないので、防御物質量の環境応答についてはGCB仮説とCNB仮説によれば同じ結果が予測されます。上で紹介したKorichevaらのメタ解析ではCNB仮説とGDB仮説はいっしょくたにされていて、同様の結論(防御に使える炭素の量を説明できるが、個々の物質の量までは説明できない)を導いています。
私はあまりこの説が好きではありません。というのは、生理学的な背景がほとんどない仮説であるため、どういうときにどういう応答がこの仮説から「予測」されるのかがよくわからないからです。仮説というよりは結果を理解するための「解釈」でしかないような気がするのですが。Korichca (2002) はCNB仮説を棄却しよう、という論争の中で、「CNB仮説を棄却してもGDB仮説があるから困らない」みたいなことを言ってますが、本当かなあ?
仮説の名前だけ聞くと、植物が環境によって防御物質の量を変えているとかいう話(つまり種内変異)に思えますが、実際には防御物質量の種間差を説明するための仮説です(Coley et al. 1985)。
植物の成長のポテンシャル(具体的には、最適な条件で育てたときの相対成長速度)は種によって違います。この違いはその植物の生育地(habitat)と相関があり、おおざっぱに言って資源(光・栄養・水など)が不足しがちな生育地にはポテンシャルが低い植物が分布しています。Coleyらは成長速度のポテンシャルが異なれば、最適な防御戦略が異なるのではないかと考えました。アイディアは以下の簡単なモデルで表されます。
成長速度=最大成長速度×(1-防御物質への投資比率)-(被食圧のポテンシャル-防御物質による回避)
ここで、「最大成長速度」は植物が防御物質へ全くバイオマスを投資せず、かつ被食されないことを仮定した成長速度です。「防御物質への投資比率」はそのままの意です。防御物質は直接は光合成や成長には正の貢献がないので、比率が大きくなれば実現される成長速度は下がります。「被食圧のポテンシャル」は、防御物質を持たなかった場合に食べられることによる成長速度の低下を意味します。「防御物質による回避」は、防御物質の濃度が高まると食べられにくくなるので被食圧が低下する様を表しています。このモデルを計算してみると、成長速度を最大にする最適な防御物質への投資比率が最大成長速度の高低によって異なることがわかります。最大成長速度が高い植物は防御物質を少なく持ったほうがよく、逆に最大成長速度が低い植物は防御物質を多く持ったほうがよいと予測されます。どうしてこのモデルではこういう結果が出るかというと、最大成長速度が高いと、そもそも食われる量が相対的に小さいので、防御物質を多くすることのメリットが小さいからです。
実際の植物では、成長速度が高い種ほど防御物質の濃度が低く、食べられやすいという結果が多く示されており(例えばColey 1988)、予測は支持されていると言えるようです。
私が一つ疑問に思ったのは、この仮説では最大成長速度が種によって違う原因をなんだと考えているのだろう?ということです。論文によれば貧資源地では、「資源を無駄遣いしないために成長速度が低いほうが有利である(成長速度が高いと器官の回転が速くなり、捨てる器官が多くてて無駄)」といった書き方をしていますが、私は「資源を無駄遣いしないために器官の寿命を長くする。そのためには器官が食べられないようするために防御への投資が多くなり、成長速度が低くなる」といったストーリーを考えていました。つまり、特定の種で成長のポテンシャルが低い理由は防御物質に多く投資しているからで、これが正しいとすると、このモデルで言う「最大成長速度」は全ての種で等しく、最適防御物質の濃度も等しいことになるんだけど・・・。
とりあえず勝手な名前をつけましたが、この手の仮説は訳しにくいのが多いです。しかも(日本人には)わかりにくい。「Apparency」ですから植物の見た目の違いによって防御のしかたが違う、という仮説のように思えますが、仮説の実体は葉が植食者に見える期間が長いか短いかによって防御のしかたが違う、という話ですから、葉寿命仮説とでも呼ぶべきかと、個人的には思います。
仮説自体はFeeny (1976) とRhoades and Cates (1976) に同時に提唱され田茂のです。葉が長寿命の植物(apparent)は、タンニンや樹脂といった、量が多く、消化されにくい防御物質をためこみ、葉が短寿命の植物(unapparent)は、アルカロイドなどの、質的な(量が少なくても効く)、あるいは毒性の防御物質を少量ためこむ、というものです。葉が長寿命の植物は、植食者に見つかりやすいので、様々な植食者が来る可能性があります。アルカロイドのような毒をため込んでも、その毒を解毒する能力を持った特殊な植食者(いわゆるspecialist)が来ると被食を防げません。そこでどのようなspecialistが来ても被食を防げるように、量的防御をするわけです。一方、葉が短寿命の植物は、植食者に見つかりにくいので、コストがかからない毒性化合物をためこむことで防御をする、という仮説です。
この仮説は「植物の防御はなぜ多様なのか」に初めて答えようとした仮説だそうで、その後の研究に与えた影響は大きかったようです。しかし、現実とは必ずしも合わないので、現在ではあまり受け入れられているわけではないようです。