岝崌惉偺惗棟惗懺妛島嵗
丂
偼偠傔偵乮200602 26乯
巜悢娭悢揑惉挿乮200011 26乯
NAR偲LAR乮200012 18乯
塰梴墫偺媧廂偺昡壙乮200705 05乯
嵟揔側抧忋晹/抧壓晹斾乮199905 27乯
丂攚宨
丂儌僨儖
惉挿夝愅墳梡僶乕僕儑儞乮200012 25乯
棯岅昞乮200101 12乯
丂怉暔偼惉挿偟傑偡丅偙偙偱偄偆惉挿偲偼丄婎杮揑偵偼姡憞廳検乮亖僶僀僆儅僗乯偑戝偒偔側傞偙偲傪堄枴偟傑偡丅堦斒偵彫偝偄屄懱偼巰偵傗偡偄偺偱丄惉挿懍搙偑掅偄傛傝偼崅偄曽偑惗懚偵偼桳棙偱偁傞偲峫偊傜傟傑偡丅偱偼偳偺傛偆側梫場偑惉挿懍搙偺崅掅傪寛傔傞偺偱偟傚偆偐丅
丂偙偙偱埖偆偺偼偄傢備傞惉挿夝愅乮growth analysis乯偱偡丅峀媊偵偼丄惉挿夝愅偲偄偆尵梩偺堄枴偡傞偲偙傠偼丄暥帤捠傝乽惉挿傪夝愅偡傞偙偲乿偲偄偆偙偲偱偡偑丄惗棟惗懺妛偺暘栰偱偼嫹媊偺乽惉挿夝愅乿偑偁偭偰丄偁傞寛傑偭偨偍嶌朄偑偁傝傑偡丅偦偆丄RGR偲偐LAR偲偐丄僔儘僂僩偵偼堄枴晄柧偺3暥帤棯岅偑懕偔悽奅偱偡丅偙傟偼偐側傝屆偔偐傜妋棫偝傟偰偄傞庤朄偱丄巚偭偨傛傝乮偲偄偭偰偼幐楃偐乯栶偵棫偪傑偡丅
丂惉挿夝愅偺杮幙偼丄惉挿懍搙偺堘偄偑壗偵桼棃偡傞偺偐丄傪偮偒偮傔傞偙偲偵偁傝傑偡丅惉挿懍搙偼懡偔偺梫場偺寢壥偱偡丅惉挿懍搙偵堘偄偑尒傜傟偨応崌丄偳偺梫場偵傛偭偰偦偺堘偄偑傕偨傜偝傟傞偐偑嫽枴偲側傞傢偗偱偡偑丄偳偺傛偆偵傾僾儘乕僠偡傞偺偑椙偄偱偟傚偆偐丅椺偊偽岝崌惉傗屇媧側偳峫偊傜傟傞梫場傪偟傜傒偮傇偟偵夝愅偡傞偺偼堦偮偺僥偱偡偑丄慱偭偨梫場偑曄傢偭偰偔傟傞偲偼昁偢偟傕尷傝傑偣傫丅梫場傪堦偮堦偮偮傇偟偰偄偒丄嵟廔揑偵惉挿偺堘偄傪夝柧偡傞偙偲傪栚巜偡偺偑儃僩儉傾僢僾側傜丄惉挿夝愅偼僩僢僾僟僂儞偺挷傋曽偱偡丅惉挿懍搙傪場悢暘夝偟偰偄偔偙偲偵傛傝丄偳偙偵堘偄偑偁傞偐傪扵偟偰偄偔曽朄偱偡丅
丂惉挿懍搙夝愅偼偳偪傜偐偲偄偆偲嶼悢偺栤戣偱偡偑丄怉暔偑偳偺傛偆偵惉挿偡傞偐傪捈娤揑偵棟夝偝偣偰偔傟傞丄傢偐傝傗偡偄棟榑揑榞慻傒偱傕偁傝傑偡丅
丂
乽偙偙偵1暘娫偵1夞暘楐偡傞嵶朎偑堦偮價儞偺拞偵擖偭偰偄傞丅1帪娫偱偙偺嵶朎偼價儞傪枮攖偵偟偰偟傑偆偲偄偆丅偱偼偙偺嵶朎偑價儞偺敿暘偺検偵側傞偺偼壗暘偐偐傞偐丠乿偲偄偆僋僀僘偑偁傝傑偟偨乮巹偑巕嫙偺崰偩偭偨偲巚偄傑偡乯丅摎偼30暘偱偼側偔丄59暘偱偡丅嵶朎偑寛傑偭偨懍搙偱暘楐偟偰偄偔丄偲偄偆偙偲偼丄帪娫t偵懳偡傞嵶朎偺悢Y偺憹偊曽偑巜悢娭悢揑偩偲偄偆偙偲偱偡丅偙偺働乕僗偩偭偨傜丄
Y = 2t
偱偡偹丅怉暔偺惉挿乮屄懱廳偺憹偊曽乯傕摨條偱偡丅怉暔偺応崌偼嵶朎偑慡偰暘楐偡傞傢偗偱偼側偄偺偱偡偑丄堦枃偺梩偺岝崌惉偵傛偭偰堦枃梩傪嶌傞偲丄惗嶻椡偑2攞偵側傝丄師偵偼2枃怴偟偔梩傪嶌傞偙偲偑偱偒傞偲偄偆傛偆偵丄暋棙揑側惉挿傪偡傞揰偵偍偄偰偼帡偨傛偆側傕偺偱偡丅偙偺偨傔丄惉挿懍搙偑帪娫偲偲傕偵憹壛偟偰偄傞偙偲偵拲堄偟偰壓偝偄丅堦斒偵丄怉暔偺惉挿偼埲壓偺幃偱昞偝傟傑偡丅
W = W0 exp(rt)
W偼屄懱廳丄W0偼帪娫0偱偺屄懱廳偲偄偄傑偡丅exp(rt)偼ert偺偙偲偱偡丅r偼學悢偱丄偙偺悢帤偺戝彫偑屄懱惉挿偺懍偄抶偄傪寛傔傑偡丅
丂r偼乽憡懳惉挿懍搙乮relative growth rate偱RGR偁傞偄偼specific growth rate乯乿偲屇偽傟傑偡丅憡懳惉挿懍搙偲偼丄惉挿懍搙乮growth rate偱GR乯傪屄懱廳偱妱偭偨傕偺偱偡丅偳偆偟偰r偑憡懳惉挿懍搙偵側傞偺偐傪愢柧偟傑偟傚偆丅屄懱偺惉挿懍搙乮扨埵帪娫摉偨傝偺惉挿検乯偼W傪t偱旝暘偡傞偙偲偵傛偭偰摼傜傟傑偡丅
dW/dt = r W0 exp(rt)
W = W0 exp(rt) 傪戙擖偟偰傗傞偲
dW/dt = r W
r = (dW/dt)/W
偲偄偆偙偲偱偡丅r偑堦掕偱偁偭偰傕屄懱僒僀僘偵傛偭偰惉挿懍搙偼曄傢偭偰偟傑偆偺偱丄r乮RGR乯偑乽惉挿偺懍偝乿偺巜昗偲偟偰巊傢傟傑偡丅
丂偝偰丄幚嵺偺怉暔偼巜悢娭悢揑側惉挿傪偡傞偺偱偡偑丄r偑偢偭偲堦掕偲偄偆偺偼側偐側偐側偔丄偪傚偭偲偯偮r偑掅壓偡傞偙偲偑懡偄偱偡丅幚尡揑尋媶偱偼丄悢擔偁傞偄偼悢廡娫偲偄偆抁偄婜娫偵尷偭偰怉暔傪惉挿偝偣丄偦偺婜娫拞偼巜悢娭悢揑惉挿傪偟偰偄傞丄偲壖掕偡傞偙偲偑懡偄偱偡丅偙偺婜娫拞偺屄懱廳偺憹壛検傪惉挿懍搙偲偟丄乽暯嬒偺屄懱廳乿偱妱傞偙偲偵傛偭偰憡懳惉挿懍搙傪寁嶼偟傑偡丅
丂傑偢丄惉挿懍搙偼埲壓偺幃偱昞偝傟傑偡丅
GR = (W2 - W1)/(t2 - t1)
W1偼堦夞栚偺應掕偺偲偒偺屄懱廳丄t1偼堦夞栚偺應掕偺偲偒偺帪娫偱偡丅憡懳惉挿懍搙RGR偼埲壓偺幃偱媮傔傞偙偲偑偱偒傑偡丅
RGR = {ln(W2) - ln(W1)}/(t2 - t1)
ln()偼帺慠懳悢偱偡丅偙傟偼偳偆傗偭偰摫偐傟傞偐偲偄偆偲丄
W2 = W1 exp(r{t2 - t1})
W2/W1 = exp(-r{t2 - t1})
ln(W2) - ln(W1) = r(t2 - t1)
偱偡丅傛偆偡傞偵丄RGR偼W傪懳悢僾儘僢僩偟偨偲偒偺捈慄偺孹偒丄偲偄偆偙偲偱偡偹丅暯嬒偺屄懱廳偼丄憡壛暯嬒偱偼側偔丄埲壓偺幃偱昞偝傟傑偡丅
Wmean = (W2 - W1)/{ln(W2) - ln(W1)}
偙偺Wmean偑壗傪堄枴偡傞偺偐丄偱偡偑丄dW/dt偑GR偲摨偠抣偵側傞偲偒偺W偲偍峫偊壓偝偄丅
丂偪側傒偵丄幚嵺偺怉暔偱RGR偑帪娫偲偲傕偵掅壓偡傞棟桼偱偡偑丄帺屓旐堿偑婲偙傞偐傜乮偮傑傝丄梩傪懡偔嶌傞偲忋偺梩偑壓偺梩傪旐堿偟丄壓偺梩偺岝崌惉検偑彮側偔側傞偐傜乯偩偲巚偭偰偄傞曽偑懡偄傛偆偱偡丅偟偐偟丄偨偄偰偄偺応崌偼塰梴墫晄懌偱婲偙傝傑偡丅杮摉偵塰梴墫嫙媼傪廩暘偵峴偊偽丄屄懱惉挿偼偐側傝挿偄帪娫巜悢娭悢揑惉挿傪堐帩偱偒傑偡丅傕偪傠傫丄偢偭偲懕偗偰偄傟偽帺屓旐堿偑婲偙偭偰偟傑偄傑偡偑丅偮偄偱偵尵偆偲丄懡偔偺尋媶偱乽塰梴墫偼廩暘梌偊偨乿偲娙扨偵偺偨傑偆恖偑懡偄偺偱偡偑丄晛捠偺幚尡偱巊偆傛偆側忦審偱偼丄偐側傝塰梴墫傪梌偊偰傕丄乽廩暘乿側偺偼敪夎捈屻丄屄懱僒僀僘偑彫偝偄偲偒偔傜偄偱偡丅屄懱偑戝偒偔側偭偰傕塰梴墫傪廩暘梌偊傛偆偲巚偭偨傜丄怉暔傪悈峩偱堢偰偰丄忢帪悈峩塼傪悅傟棳偟懕偗傞乮free access偲偄偆乯偔傜偄偺偙偲偼偟側偄偲偄偗傑偣傫丅
丂
偝偰丄偁側偨偺慜偵RGR偑堎側傞擇偮偺怉暔偑偁偭偨偲偟傑偡丅偁側偨偼摉慠偳偆偟偰偙偺惉挿懍搙偺堘偄偑婲偙傞偺偐丄偦偺儊僇僯僘儉傪抦傝偨偄偲巚偆偼偢偱偡丅傁偭偲巚偄偮偔偺偼乽岝崌惉懍搙偺堘偄乿偱偟傚偆丅岝崌惉傪偨偔偝傫偟偰偄傞偐傜惉挿偑懍偄傢偗偱偡偐傜偹丅尵偄姺偊傟偽丄RGR偭偰偺偼屄懱廳摉偨傝偺岝崌惉懍搙傪尒偰偄傞傛偆側傕偺偱偡偐傜乮尩枾偵偼屇媧懍搙傕廳梫偱偡偑乯丅偟偐偟偦傟偱偼乽惉挿乿傪乽岝崌惉乿偵尵偄姺偊偰偄傞偩偗偱偡丅傕偆彮偟峫偊傑偟傚偆丅
丂堦斒揑偵尵偭偰丄岝崌惉丄偲偄偆偺偼梩偑偡傞傕偺偱偡乮椺奜偼偁傝傑偡偑乯丅偟偨偑偭偰梩傪偨偔偝傫帩偭偰偄傞怉暔偺RGR偑崅偦偆側婥偑偟傑偡丅偄傗偄傗偦偆偱偼側偔丄屄乆偺梩偺岝崌惉懍搙偑崅偄怉暔偺RGR偑崅偦偆側婥傕偟傑偡丅偙傟傪悢幃偱昞偟偰傒傑偟傚偆丅
RGR = GR/W = GR/LA 亊 LA/W
偙偙偱丄LA偼屄懱偑帩偮梩柺愊偱偡丅塃曈偺戞堦崁偼乽梩柺愊偁偨傝偺惉挿懍搙乿傪昞偟傑偡丅偙傟偼丄梩柺愊偁偨傝偳傟偩偗偺摨壔嶻暔傪摼偰偄傞偐偱丄偮傑傝丄梩柺愊偁偨傝偺岝崌惉懍搙傪斀塮偟偰偄傞偲峫偊傜傟偰偄傑偡丅戞擇崁偼乽屄懱廳偁偨傝偺梩柺愊乿偱丄偳傟偩偗懡偔梩傪帩偭偰偄傞偐乮梩柺愊傪揥奐偟偰偄傞偐乯偺巜昗偵側傝傑偡丅傕偟梩傪偨偔偝傫帩偭偰偄傞偙偲偵傛偭偰RGR偑崅偄偺側傜LA/W偑崅偔丄梩偺岝崌惉懍搙偑崅偄偙偲偵傛偭偰RGR偑崅偄偺側傜GR/LA偑崅偄偼偢偱偡丅LA/W傪Leaf Area Ratio乮棯偟偰LAR乯丄GR/LA傪Net Assimilation Rate乮棯偟偰NAR乯偲屇傫偱偄傑偡丅擔杮岅栿偱偡偑丄NAR偼乽弮摨壔懍搙乿偲栿偣傑偡偑丄LAR偼擄偟偄偱偡偹丅屆偄嫵壢彂偵偼乽梩柺愊斾乿偲捈栿偝傟偰偄傞偙偲偑懡偄偲巚偄傑偡丅
丂NAR偲LAR偵暘妱偟偨偺偼旕忢偵戝偒側堄枴偑偁傝傑偡丅偦傟偼丄RGR偑乽惗棟妛揑摿惈乿偲乽宍懺妛揑摿惈乿偺擇偮偺塭嬁傪庴偗傞偙偲丄偦偟偰丄偦傟傜傪暘棧偟偰夝愅偡傞偙偲偑偱偒傞偙偲偱偡丅偨偩丄NAR偼乽梩偺岝崌惉亅屄懱偺屇媧乿傪梩柺愊偱妱偭偰偄傞偺偱丄尩枾偵偼岝崌惉懍搙偦偺傑傑偲偼尵偊傑偣傫丅偨偩丄偙偺夝愅傪偡傟偽RGR偺堘偄傪妋幚偵偳偪傜偐偵婣偡傞偙偲偑偱偒傞丄偲偄偆揰偱丄扨側傞岝崌惉懍搙偺應掕偲偼堘偆堄枴偑偁傝傑偡丅
丂偝偰LAR偼宍懺妛揑摿惈傪斀塮偟偰偄傞丄偲彂偒傑偟偨偑丄LAR帺懱偼傗傗傢偐傝偵偔偄僷儔儊乕僞偱偡丅梩柺愊偲偄偆2師尦偺僷儔儊乕僞傪屄懱廳偲偄偆3師尦偺僷儔儊乕僞偱妱偭偰偄傞傢偗偱偡偐傜丅LAR傪憹傗偡偨傔偵丄偳偆偄偆偙偲偑偱偒傞偐傪峫偊偰傒傑偟傚偆丅
丂LAR傪憹傗偡曽朄偼擇偮峫偊傜傟傑偡丅堦偮偼丄僶僀僆儅僗傪梩偵懡偔夞偡偙偲偱偡丅塰梴惉挿婜側傜偽僶僀僆儅僗偼梩丒宻丒崻偺偄偢傟偐偵夞偝傟偰偄傑偡丅偙偺偆偪梩偵懡偔傑傢偣偽丄LAR偼憹偊傑偡丅傕偆堦偮暿偺曽朄偑偁傝傑偡丅偦傟偼丄梩傪敄偔偟偰梩柺愊傪峀偘傞偙偲偱偡丅偙傟傕傑偨悢幃偵偟偰傒傑偟傚偆丅
LAR = LA/W = LA/LW 亊 LW/W
塃曈戞堦崁偼梩柺愊傪梩廳偱妱偭偨傕偺偱偡丅偙偺抣偑崅偄偲偄偆偙偲偼丄梩偵搳帒偝傟偨僶僀僆儅僗偱傛傝峀偄梩傪嶌偭偰偄傞偲偄偆偙偲傪堄枴偟傑偡丅Specific Leaf Area乮棯偟偰SLA乯偲尵偄傑偡丅擔杮岅栿偼梩柺愊斾廳偩偭偨偐側偁丅偪側傒偵丄偙偙偱偺乽specific乿偺堄枴偼乽廳偝偁偨傝乿偲偄偆偙偲偱偡丅塃曈戞擇崁偼梩廳傪屄懱廳偱妱偭偨傕偺偱丄屄懱廳偺偆偪梩廳偺妱崌傪帵偟傑偡丅婍姱娫偺暔幙暘攝傪斀塮偟偰偄傞傢偗偱偡丅Leaf Wight Ratio乮LWR乯偁傞偄偼Leaf Mass Ratio乮LMR乯偲尵偄傑偡丅嵟嬤偼LMR偺曽偑懡偄偱偡偹丅擔杮岅栿偼丒丒丒梩廳斾偩偭偨偲巚偄傑偡丅帤偩偗偱偼傛偔傢偐傜側偄偱偡偑丅
丂偲偄偆傢偗偱丄
RGR = NAR 亊 LAR = NAR 亊 LMR 亊 SLA
偲偄偭偨僷儔儊乕僞偱RGR偺堘偄傪夝愅偡傞偙偲偑偱偒傑偡丅扨側傞偐偗嶼偵尒偊傑偡偟丄幚嵺偦偆側偺偱偡偑丄偨偩傗傒偔傕偵偐偗嶼偵偟偰偄傞偺偱偼側偔丄惗暔妛揑偵堄枴偺偁傞夝愅傪偡傞偨傔偺偐偗嶼偩偲峫偊偰壓偝偄丅
丂偪側傒偵丄SLA偺媡悢丄偡側傢偪丄梩柺愊偁偨傝偺梩廳傪Leaf Mass Area乮LMA乯偲尵偄傑偡丅巹偼拏慺傗傜岝崌惉傗傜梩柺愊偁偨傝偱暔幙娷検傪昞偡偙偲偑懡偄偺偱丄SLA傛傝偼LMA傪巊偆偙偲偑懡偄偱偡丅側偍丄LMA偼偐偮偰偼Specific Leaf Weight乮SLW乯偲屇偽傟偰偄傑偟偨丅偟偐偟丄忋偵弎傋偨傛偆偵Specific偼乽廳偝偁偨傝乿偺堄枴側偺偱丄SLW偼乽廳偝偁偨傝偺廳偝乿偲偄偆偙偲偵側傝丄忢偵1偱偡丅偲偄偆偙偲偱丄嵟嬤偼巊傢側偄偙偲偑悇彠偝傟偰偄傑偡乮榑暥偵彂偔偲愨懳僋儗乕儉偑偮偒傑偡乯丅LMA傪巊偆傛偆偵偟傑偟傚偆丅
丂
怉暔偺惉挿偵偼岝崌惉偑嵟傕廳梫側梫慺偱偡偑丄岝崌惉偩偗偱偼惉挿偼偱偒傑偣傫丅悈傗拏慺丒儕儞偲偄偭偨懠偺塰梴墫傕晄壜寚偱偡丅偙傟傜偺帒尮偼庡偵崻偱媧廂偝傟傑偡丅崻偺婡擻偲惉挿偺娭楢傪挷傋傞偨傔偵崻偺惈幙偺悢抣壔偑峴傢傟偰偄傑偡丅杮棃偼崻偺婡擻偵偮偄偰偩偗偱堦嶜杮偑彂偗傞偔傜偄偺帠暱偑偁傞偺偱偡偑丄偁傑傝傛偔抦傝傑偣傫偟丄偙偙偱偼惉挿夝愅偱巊傢傟傞庡側僷儔儊乕僞傪徯夘偡傞偵棷傔傑偡丅
丂惉挿夝愅偲偺娭楢偱偼拏慺偵拝栚偡傞偙偲偑懡偄偱偡丅懠偺尦慺偑廳梫偱側偄偲偄偆傢偗偱偼側偄偱偡偑丅昿斏偵巊傢傟傞僷儔儊乕僞偼崻偺廳検偁偨傝偺拏慺媧廂懍搙乮specific absorption rate, SAR乯偱偡丅偙傟偼扨埵帪娫偁偨傝偵怉暔屄懱偑媧廂偟偨拏慺傪崻廳検偱妱偭偨傕偺偱偡丅
丂SAR偼丄惉挿夝愅儌僨儖偱偼拏慺塰梴忦審偺巜昗偲偟偰巊傢傟傞偙偲偑懡偄偱偡丅SAR偑崅偄亖塰梴忦審偑椙偄丄偲偄偆傛偆偵峫偊傞傢偗偱偡丅偨偩丄摨偠塰梴忦審偱傕屄懱僒僀僘傗庬偵傛偭偰堎側傝傑偡丅條乆側梫場偑SAR偺堘偄偵塭嬁偟傑偡丅堦偮偼崻偺宍懺偱偁傞偲峫偊傜傟偰偄傑偡丅扨埵廳検偁偨傝挿偄丄偁傞偄偼戝偒側昞柺愊傪傕偮崻偼搚忞偲偺愙怗柺愊偑戝偒偄偨傔懡偔偺拏慺傪媧廂偱偒傞偩傠偆丄偲偄偆傕偺偱偡丅偙偺傛偆側宍懺偺堘偄傪悢抣壔偡傞偨傔偵崻廳偁偨傝偺崻挿乮specific root length, SRL乯傗崻廳偁偨傝偺崻昞柺愊偑巊傢傟傑偡丅崻昞柺愊偼側偐側偐應傝偯傜偄偺偱丄SRL偑巊傢傟傞偙偲偑懡偄傛偆偵巚偄傑偡丅
丂
丂偙偙偱偼丄惉挿夝愅儌僨儖傪墳梡偟偨夝愅傪徯夘偟傑偡丅
丂塰梴傗岝側偳偺惗堢忦審偑曄壔偡傞偲丄屄懱撪偺婍姱娫偺僶僀僆儅僗暘攝偑曄壔偟傑偡丅摿偵丄抧忋晹/抧壓晹偺斾偑曄壔偡傞偙偲偑傛偔抦傜傟偰偄傑偡乮抶偔偲傕1930擭戙偵偼抦傜傟偰偄偨傛偆偱偡)丅抧忋晹偺僶僀僆儅僗偼扽慺傪妉摼偡傞偨傔丄抧壓晹偺僶僀僆儅僗偼拏慺傪妉摼偡傞偨傔偵昁梫偱偡丅拏慺偑懌傝側偄傛偆側忦審偱偼丄抧壓晹傊偺僶僀僆儅僗偺暘攝傪懡偔偟偰傛傝拏慺偑媧廂偱偒傞傛偆偵偟丄岝偑懌傝側偄忦審偱偼抧忋晹傊偺搳帒傪懡偔偟偰扽慺傪傛傝媧廂偱偒傞傛偆偵偡傞偲峫偊傞偲丄抧忋晹/抧壓晹斾偺曄壔偼尰幚偺曄壔傪傛偔愢柧偟傑偡丅偙傟偼旕忢偵棟夝偟傗偡偄榑棟偱偡丅偟偐偟丄娙扨側悢棟儌僨儖傪嶌偭偰傒傞偲丄偙偺扨弮側榑棟偑堄奜偵惓偟偔側偄丄偲偄偆偐丄尰幚揑側梊應傪偡傞偨傔偵偼偄偔偮偐偺壖掕乮傕偟偔偼惂栺乯傪庢傝擖傟傞偙偲偑昁梫偱偁傞偙偲偵婥偯偒傑偡丅偙偙偱偼丄嵟揔側抧忋晹/抧壓晹斾傪峫偊傞偵偁偨偭偰丄峫椂偟側偗傟偽偄偗側偄梫場偵偮偄偰彂偒傑偡丅
丂
丂1960擭戙偵偼丄乽抧忋晹偲抧壓晹偺妶惈乮扽慺偲拏慺偺妉摼乯偺僶儔儞僗傪偲傞乿偲偄偆丄屻悽偺儌僨儖偵斾傋傞偲丄敊慠偲偟偨傾僀僨傿傾丒儌僨儖偑偁偭偨傛偆偱偡乮婡擻暯峵儌僨儖丄functional equiribrium; Brouwer 1962; Davidson 1969; Thornley 1972乯丅Davidson (1969) 偼抧忋晹偺扽慺媧廂偲抧壓晹偺拏慺媧廂偑摨偠偵側傞傛偆偵挷愡偝傟偰偄傞偺偩偲峫偊傑偟偨丅Thornley (1972) 偼丄屄懱偺扽慺/拏慺斾傪堦掕偵偡傞傛偆側抧忋晹/抧壓晹斾傪儌僨儖偱帵偟傑偟偨丅偟偐偟丄幚嵺偵偼扽慺/拏慺斾偼堦掕偵側傞傢偗偱偼側偄偺偱丄尰幚傪愢柧偡傞偵偼晄廩暘側儌僨儖偱偟偨丅
丂80擭戙偵側傞偲丄乽揔墳搙傗憡懳惉挿懍搙乮RGR乯傪嵟戝偵偡傞抧忋晹/抧壓晹斾乿偑峫偊傜傟傞傛偆偵側偭偰偒傑偟偨乮Iwasa and Roughgarden 1984; Hunt and Nicholls 1986; Johnson and Thornley 1987乯丅偙傟傜偺儌僨儖偑偳偆偄偆巇慻傒偵側偭偰偄傞偺偐傛偔偼抦傝傑偣傫偑丄Hilbert (1989) 偵傛傟偽丄偙傟傜偺儌僨儖偱偼丄乽抧忋晹偺扽慺妉摼擻椡偑抧忋晹拏慺娷検偵埶懚偡傞乿偲偄偆壖掕偑側偄偺偩偦偆偱偡丅摨偠抧忋晹僶僀僆儅僗傪帩偭偰偄偰傕丄拏慺娷検偑彮側偗傟偽岝崌惉擻椡偼掅偄偼偢偱丄偙偺壖掕傪擖傟側偗傟偽崻偺栶妱偺昡壙傪岆傞偙偲偑梊憐偱偒傑偡丅
丂堦曽丄Ingestad偲Agren偺僌儖乕僾偼丄nitrogen productivity偲偄偆奣擮偺傕偲偱抧忋晹/抧壓晹斾傪峫嶡偟傑偟偨丅怉暔傪條乆側塰梴忦審偱惗堢偝偣傞偲丄屄懱偺憡懳惉挿懍搙偲拏慺娷検偑惓斾椺偡傞偙偲偑抦傜傟偰偄傑偡乮Ingestad 1979乯丅偮傑傝偙傟偼曐桳拏慺偁偨傝偺惉挿懍搙乮偙傟偑nitrogen productivity乯偑堦掕偱偁傞偙偲傪堄枴偟傑偡丅Ingestad傜偼nitrogen productivity堦掕傪朄懃偲峫偊丄偙傟傪幚尰偡傞偨傔偵偼抧忋晹/抧壓晹斾偑偳偺傛偆偵曄壔偡傞偺偐傪峫嶡偟偨傛偆偱偡乮Agren and Ingestad 1987乯丅偙偺峫偊曽偑懨摉偐斲偐偼偁偲偱愢柧偟傑偡丅
丂Hirose (1988) 傗Hilbert (1989) 偺儌僨儖偱偼丄乽扽慺偼抧忋晹偵傛偭偰妉摼偝傟傞乿乽拏慺偼抧壓晹偵傛偭偰妉摼偝傟傞乿偺懠偵丄乽抧忋晹偁偨傝偺扽慺妉摼懍搙偼抧忋晹偺拏慺擹搙偵埶懚偡傞乿偲偄偆壖掕偑慻傒崬傑傟偰偄傑偡丅偱偼丄偙傟傜偺壖掕偐傜乽嵟揔側抧忋晹/抧壓晹斾乿偼偳偺傛偆偵寁嶼偝傟傞偱偟傚偆偐丠
丂
偙偙偱偼丄乽嵟揔側抧忋晹/抧壓晹斾乿偵偼傛偗偄側壖掕偑堄奜偵昁梫偩丄偲偄偆偙偲傪棟夝偟偰偄偨偩偔偨傔丄嵟弶偵嵟傕扨弮側儌僨儖傪嶌傝丄彊乆偵尰幚揑側儌僨儖傪峔抸偟偰偄偔偲偄偆丄乽巚峫幚尡偺幚尡乿傪偟偰傒傑偡丅
丂
戞侾儌僨儖
偱偼丄乽扽慺偼抧忋晹偵傛偭偰妉摼偝傟傞乿乽拏慺偼抧壓晹偵傛偭偰妉摼偝傟傞乿乽抧忋晹偁偨傝偺扽慺妉摼懍搙偼抧忋晹偺拏慺擹搙偵埶懚偡傞乿偲偄偆俁偮偺壖掕傪幃偵偟偰傒傑偟傚偆丅
丂偙偙偱偼丄娙扨偵偡傞偨傔偵丄宻偺懚嵼偼柍帇偟傑偟傚偆乮儘僛僢僩怉暔傪峫偊傞偲傛偄乯丅梩=抧忋晹偲偟傑偡丅
丂惉挿懍搙乮dW/dt乯偼抧忋晹偺扽慺妉摼偵傛傝傕偨傜偝傟傑偡偐傜丄埲壓偺幃偱昞偡偙偲偑偱偒傑偡丅
dW/dt = A LW (1)
偙偙偱A偑梩廳検偁偨傝偺暔幙惗嶻検乮NAR亊SLA偵憡摉乯偱丄LW偑梩廳検偱偡丅憡懳惉挿懍搙偼椉曈傪屄懱廳乮W乯偱妱傞偙偲偵傛傝摼傜傟傑偡丅
(1/W)dW/dt = A LMR (2)
偙偙偱丄LMR偼梩廳/屄懱廳斾乮LW/W乯偱偡丅
丂師偵丄屄懱偺拏慺媧廂懍搙dN/dt傪掕媊偟傑偟傚偆丅
dN/dt = SAR Wr = SAR fr W = SAR (1-fs)W (3)
偙偙偱SAR偑崻廳偁偨傝偺拏慺媧廂検丄RM偑崻廳偱偡丅SAR偼怉暔偼挷愡偱偒側偄偲壖掕偟丄偙偺抣偑塰梴忦審偺巜昗偲側傝傑偡乮昻塰梴忦審傎偳SAR偑彫偝偄乯丅
丂梩廳検偁偨傝偺暔幙惗嶻検A偑抧忋晹偺拏慺擹搙偵埶懚偡傞偲偄偆壖掕傪擖傟偰傒傑偟傚偆丅傑偢丄嵟傕扨弮偵丄抧忋晹偺拏慺擹搙偲A偑惓斾椺偡傞偲偄偆壖掕偵偟偰傒傑偟傚偆丅
A = C NL/LW (4)
NL偼抧忋晹偑帩偮拏慺偺愨懳検偱丄C偼斾椺掕悢偱偡丅偙偙偱丄榖傪偝傜偵扨弮偵偡傞偨傔偵丄抧忋晹偺拏慺擹搙偲抧壓晹偺拏慺擹搙偑摨偠偲偟傑偟傚偆乮幚嵺偼抧忋晹偺傎偆偑偼傞偐偵崅偄亅徻嵶屻弎乯丅
NL/LW = N/W (5)
偱偼丄暯峵忬懺傪壖掕偟[N/W = (dN/dt)/(dW/dt)]丄偙傟傜偺幃傪慻傒崌傢偣偰傒傑偟傚偆丅埲壓偺幃偵側傞偼偢偱偡丅
[(1/W)dW/dt]2 = C SAR (1-LMR)LMR (6)
偙偙偱丄RGR傪嵟戝偵偡傞LMR傪媮傔傑偟傚偆丅偙偺幃偺僷儔儊乕僞偼慡偰惓偺抣傪庢傝傑偡偐傜丄塃曈偼忋偵撌偺擇師嬋慄偵側傞偙偲偑傢偐傝傑偡丅RGR偺擇忔傪嵟戝壔偡傞偺傕RGR傪嵟戝壔偡傞偺傕摨偠偱偡偐傜丄偙偺幃偺旝暘抣偑0偵側傞抣傪寁嶼偡傞偲丄夝偼丄
LMR = 0.5
偲側傝傑偡丅偙偺夝偺堄枴偡傞偲偙傠偼丄乽嵟揔側抧忋晹/抧壓晹斾偼塰梴忦審乮SAR偺偙偲乯偵傕C偵傕埶懚偣偢0.5偱堦掕偱偁傞乿偲偄偆偙偲偱偡丅偳偆偟偰偦偆側傞偺偐傪捈娤揑偵愢柧偡傞偺偼擄偟偄偺偱偡偑丄偙偙偵嫇偘偨壖掕偱偼丄LMR傪0.5偐傜曄偊偰傕丄曄偊偨暘偵憡摉偡傞A偺曄壔偑摼傜傟側偄丄偲偄偆偙偲偵傛傝傑偡丅偲偄偆傢偗偱丄乽扽慺偼抧忋晹偵傛偭偰妉摼偝傟傞乿乽拏慺偼抧壓晹偵傛偭偰妉摼偝傟傞乿乽抧忋晹偁偨傝偺扽慺妉摼懍搙偼抧忋晹偺拏慺擹搙偵埶懚偡傞乿偲敊慠偲峫偊偨偩偗偱偼抧忋晹/抧壓晹斾偺曄壔偼愢柧偱偒側偄丄偲偄偆偙偲偑傢偐偭偰偄偨偩偗偨偱偟傚偆偐丅
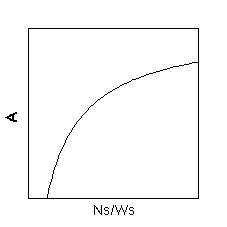
丂偱偼丄偙傟偵偳偆偄偆壖掕傪壛偊傞偲抧忋晹/抧壓晹斾偺曄壔偑愢柧偱偒傞偱偟傚偆偐丠丂摎偼丄乽抧忋晹偁偨傝偺扽慺妉摼懍搙偲抧忋晹偺拏慺擹搙偺娭學偼丄X愗曅偑惓偺抣傪帩偮嬋慄偱偁傞乿偲偄偆壖掕偱偡丅椺偊偽埲壓偺恾偺傛偆側姶偠偱偡丅Ns/Ws偼梩偺拏慺擹搙乮NL/LW乯偺偙偲偱偡丅
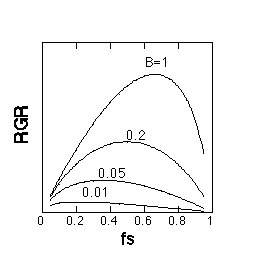
偙偺壖掕傪擖傟傞偲丄埲壓偺恾偺傛偆偵塰梴忦審乮SAR乯偑崅偄傎偳抧忋晹傊偺僶僀僆儅僗搳帒偑戝偒偔側傞丄偲偄偆夝傪摼傞偙偲偑偱偒傑偡丅壓偺恾偱B偼SAR丄fs偼LMR偺偙偲偱偡丅
偲偄偆傢偗偱丄惓偟偄乽嵟揔側抧忋晹/抧壓晹斾乿偺梊應偵偼丄乽抧忋晹偁偨傝偺扽慺妉摼懍搙偲抧忋晹偺拏慺擹搙偺娭學乿傪抦傞偙偲偑昁梫偱偁傞丄偲偄偆偙偲傪偙偙偱偼棟夝偟偰壓偝偄丅
丂偪側傒偵丄嵟揔側fs偼傕偭偲娙扨偵夝偔偙偲偑偱偒傑偡丅A偩偗偱側偔丄LMR傗RMR (崻廳/屄懱廳斾乯傕拏慺擹搙n(= N/W)偺娭悢偵側傞偼偢偱偡偐傜丄幃(2)傪巊偭偰丄RGR偺旝暘抣偑0偵側傞傛偆側曽掱幃傪夝偔偲丄埲壓偺夝偑摼傜傟傑偡丅
LMR dA/dn = -A dLMR/dn
偙偺幃傪枮懌偝偣傞fs偑嵟揔側LMR偲偄偆偙偲偵側傝傑偡乮Hilbert 1989乯丅
丂
戞俀儌僨儖
戞侾儌僨儖偱偼丄抧忋晹偺暔幙惗嶻懍搙偲拏慺擹搙傪丄偦傟偧傟廳検偁偨傝偱峫偊偰偄傑偟偨丅偟偐偟丄偙偺乽岝崌惉偺惗棟惗懺妛島嵗乿偱偼岝崌惉懍搙傗拏慺娷検偼梩柺愊偁偨傝偱峫偊偰偒傑偟偨丅廳検偁偨傝偱峫偊傞偙偲偲柺愊偁偨傝偱峫偊傞偙偲偲偺堘偄偵偮偄偰偼條乆側媍榑偑偁傝傑偡偑丄庴岝偺廳梫惈傪峫偊傞偲丄傗偼傝暔幙惗嶻偼梩柺愊偁偨傝偱峫偊傞傋偒偱偡丅
丂偦偙偱丄崱搙偼梩柺愊偺奣擮傪擖傟偰傒傑偟傚偆丅忋摨條丄抧忋晹=梩偲偡傞偲丄
LA = LW/LMA
Narea = Nmass亊LMA
偲側傝傑偡丅偙偙偱丄LA偑梩柺愊丄LMA偑梩柺愊偁偨傝偺梩廳乮SLA偺媡悢乯丄Nmass偑梩廳検偁偨傝偺拏慺娷検丄Narea偑梩柺愊偁偨傝偺拏慺娷検偲側傝傑偡丅偦偟偰丄梩柺愊偁偨傝偺暔幙惗嶻検NAR偑Narea偲埲壓偺娭學偵偁傞偲偟傑偟傚偆丅
NAR = C(Narea-k)/(Narea+h)
k偲h偼掕悢偱偡丅Aarea偼偄偆側傟偽堦擔偺岝崌惉検偵偁偨傝傑偡乮偙偺幃偼丄忋偺恾偺A偲Ns偺幃偲摨偠娭悢傪巊偭偰偄傑偡乯丅RGR偼埲壓偺傛偆偵昞偝傟傑偡丅
(1/W)dW/dt = NAR/LMA亊LMR
偙偙偐傜丄RGR傪嵟戝偵偡傞LMA偲fs偺慻傒崌傢偣傪寁嶼偡傞偙偲偵側傝傑偡丅偲偄偭偰傕丄偙偪傜偼戞侾儌僨儖偺傛偆偵娙扨偵偼夝偗傑偣傫偺偱乮夝偗傞偐傕偟傟側偄偗偳巹偵偼傢偐傜側偄乯丄僔儈儏儗乕僔儑儞傪傗偭偨摎傪弎傋傑偡丅嵟揔夝偼丄昻塰梴傎偳LMR偑彫偝偔丄LMA偑崅偔側傞丄偲偄偆傕偺偱偡丅偙偙偵偼傕偆堦偮忦審偑偁偭偰丄LMA偼Narea傪堦掕偵偡傞傛偆偵曄壔偟傑偡丅Narea偼塰梴忦審偵埶懚偣偢丄堦掕偱偁傞丄偲偄偆夝偑弌傞傢偗偱偡丅
丂Narea偑塰梴忦審偵埶懚偟側偄偲偄偆偺偼丄尰幚偺怉暔偺娤嶡偲偼柕弬偟偰偄傑偡丅偙傟偼幚偼婛偵堦搙乽嵟揔側拏慺暘攝乿偱愢柧偟偰偄傞偙偲偱偡丅偙偺儌僨儖偱偼丄NAR偑Narea偵偺傒埶懚偟偰偄傞偨傔丄偙偆偄偆摎偑弌偰偒偰偟傑偆偲傕偄偊傑偡丅懠偵丄側傫傜偐偺惂栺偑偁傞偙偲傑偱峫偊側偄偲丄Narea偑塰梴忦審偵傛偭偰堎側傞偙偲傪愢柧偱偒傑偣傫丅乽昻塰梴偩偐傜拏慺偑懌傝側偔偰Narea偑壓偑傞乿偲偄偆愢柧偼丄幚偼丄棟榑揑偵偼偐側傝晄廩暘側偺偱偡丅
丂偱偼丄偙傟傑偱偺尋媶偱偼偙偺乽側傫傜偐偺惂栺乿傪偄偭偨偄壗偲峫偊偰偒偨偱偟傚偆偐丠丂堦偮偺椺偼Hirose (1986) 偱偡丅偙偺巇帠偱偼丄LMA偲梩偺姡廳偁偨傝偺拏慺娷検乮Nmass乯偑屄懱偺楊傗塰梴忦審偵埶懚偟側偄偙偲傪尒弌偟丄偙偺娭學(Nmass偑崅偄偲LMA偑掅偄乯傪惂栺偲偟偰梡偄偰偄傑偡丅偙偙偱偼丄Nmass傪梌偊傞偲偙偺娭學偵傛偭偰Narea偑寛傑偭偰偟傑偆偨傔丄乽昻塰梴偩偐傜拏慺偑懌傝側偔偰Narea偑壓偑傞乿偲偄偆偺偼丄愢柧偝傟傞傕偺偱偼側偔丄帠幚忋乽壖掕乿偲偄偆偙偲偵側偭偰偟傑偄傑偡丅van der Werf et al. (1993) 偱傕摨偠壖掕偑巊傢傟偰偄傑偡丅傕偆堦偮偺椺偼丄Hilbert and Reynolds (1991) 偱偡丅偙偺巇帠偱偼丄LMA傪堦掕偲偟偰偄傑偡乮偨偩偟岝忦審偑曄傢傞偲曄傢傞乯丅偲偄偆偙偲偱丄乽塰梴忦審偵傛偭偰側偤Narea偑偽傜偮偔偺偐乿偼傑偩乽壖掕乿偲偟偰梡偄傜傟偰偄傞抜奒偱丄偳偆偟偰偦偆側傞偺偐丄偼枹偩傢偐偭偰偄側偄偲偄偆偙偲偲丄偙偺乽壖掕乿傪擖傟側偄偲丄尰幚偲偼堎側傞抧忋晹/抧壓晹斾偑寁嶼偝傟偰偟傑偆丄偲偄偆偙偲傪偙偙偱偼偛棟夝壓偝偄丅
丂
偙偺懠廳梫側偙偲
丂偙偙傑偱偺愢柧偱偼丄梩偺拏慺擹搙偲屄懱偺拏慺擹搙偑摨偠偱偁傞偲壖掕偟偰偒傑偟偨丅偟偐偟丄幚嵺偵偼梩偺拏慺擹搙偼屄懱偺拏慺擹搙傛傝傕偼傞偐偵崅偄偱偡丅偲偄偆偙偲偼丄偮傑傝丄崻傗宻偺拏慺擹搙偑掅偄丄偲偄偆偙偲偱偡丅宱尡揑偵偼丄梩丒宻丒崻偺拏慺擹搙偼斾椺娭學偵偁傝丄晛捠偺儌僨儖偱偼偙偺娭學傪梡偄偰偄傑偡乮Hirose 1988; Kachi & Rorison 1989; van der Werf et al. 1993乯丅偟偐偟丄偱偒傟偽丄宻傗崻偺拏慺擹搙偑寛傑傞昁慠惈傕抦傝偨偄傕偺偱偡丅梩偺拏慺偼丄岝崌惉偵娭學偑偁傞丄偲偄偆偙偲偱愢柧偱偒傑偡偑丄崻傗宻偺拏慺擹搙偑偳偆寛傑偭偰偄傞偐偼傑偩傛偔傢偐偭偰偄傑偣傫丅扨側傞擹搙暯峵偱愢柧偑偮偔偺偱偟傚偆偐丒丒丒丅
丂
偮傑傞偲偙傠丄GR/W = GR/LA 亊 LA/W偺傛偆偵丄偲偁傞僷儔儊乕僞乮偙偙偱偼LA乯傪娫偵嫴傫偱傗傞偙偲偵傛偭偰GR/W傪暘夝偡傞丄偲偄偆偺偑丄悢幃忋偺惉挿夝愅偺杮幙偱偡丅偙傟偼LA偱側偔偰傕偄偄傢偗偱丄椺偊偽丄GR/W = GR/(姫僸僎偺悢) 亊 姫僸僎偺悢/W偺傛偆側傢偗偺傢偐傜傫夝愅傕壜擻側傢偗偱偡丅傕偪傠傫偙偺夝愅偵惗暔妛揑側堄枴偑偁傞傛偆側婥偼偟傑偣傫偑乮徫乯丅偟偐偟丄乽惗暔妛揑偵堄枴偺偁傞乿夝愅偼懠偵傕偁傞偼偢偱偡丅
丂戙昞揑側傕偺偼丄拏慺惗嶻惈乮Nitrogen productivity丄NP乯傪梡偄偨傕偺偱偡丅NP偼GR/N丄偮傑傝屄懱偑帩偮拏慺検偁偨傝偺惉挿懍搙偱偡乮Ingestad 1979乯丅偮傑傝丄
RGR = NP 亊 PNC
GR/W = GR/N 亊 N/W
偙偙偱丄PNC偼屄懱拏慺擹搙乮Plant Nitrogen Concentration丄廳検摉偨傝偺拏慺検乯偱偡丅偙偺幃偼丄Ingestad傜偺僌儖乕僾偺堦楢偺尋媶偵傛傝抦傜傟傞傛偆偵側傝傑偟偨丅斵傜偼丄堎側傞塰梴忦審偱怉暔傪堢偰傞偲丄RGR偲PNC偺娭學偼傎傏捈慄偲側傝丄NP偼傎傏堦掕偲側傞偲偄偆宱尡懃傪尒弌偟傑偟偨丅偮傑傝丄N傪娫偵嫴傫偱傗傞偙偲偱怴偨側帇揰傪庤偵擖傟偨傢偗偱偡丅
丂忋幃偵斾傋傞偲慡慠桳柤偱偼偁傝傑偣傫偑丄嵟嬤変乆偺僌儖乕僾偱偼埲壓偺傛偆側幃傪採埬偟傑偟偨乮Hikosaka et al. 1999a乯丅
ARGR = AGR/AW = GR/F 亊 F/AW
偙偙偱丄AGR偼抧忋晹廳検偁偨傝偺抧忋晹廳検憹壛懍搙乮偮傑傝抧忋晹偩偗偱尒偨RGR乯丄AGR偼抧忋晹廳検憹壛懍搙丄AW偼抧忋晹廳検丄F偼屄懱偺岝媧廂検偱偡丅偙偺幃傪巊偭偰壗傪偟偨偐偭偨偐偲偄偆偲丄孮棊撪偱岝偺嫞憟傪偟偰偄傞屄懱偺惉挿傪夝愅偟偨偐偭偨偺偱偡丅孮棊撪偱偼壓晹傎偳埫偄偲偄偆娐嫬偵側偭偰偄傑偡丅偙偺拞偱屄懱偼尷傝偁傞岝傪偱偒傞尷傝媧廂偟傛偆偲岺晇傪偟偰偄傞偲峫偊傜傟傑偡丅偙偺岺晇傪F/W偲偟偰昞偟偰傒偨傢偗偱偡乮Hirose and Werger 1995乯丅偳偆偄偆偙偲偐偲偄偆偲丄梩偺栶妱偼岝傪媧廂偡傞偙偲偱偡丅偝傜偵丄宻偺栶妱偼梩傪巟偊傞偙偲偱偡丅扨弮偵峫偊偰丄梩偲宻偵搳帒偝傟偨僶僀僆儅僗偼岝傪媧廂偡傞偨傔偺搳帒偲尒傞偙偲偑偱偒傑偡丅偮傑傝丄F/AW偼丄F傪儀僱僼傿僢僩丄抧忋晹廳検傪僐僗僩偲偟偨丄儀僱僼傿僢僩/僐僗僩斾側傢偗偱偡乮Hirose and Werger 1995乯丅偱丄GR/F斾偼丄媧廂偱偒偨岝傪偳傟偩偗惉挿偵岠棪傛偔揮姺偱偒偨偐丄偲偄偆巜昗偵側傝傑偡丅変乆偼F/AW傪乽岝妉摼岠棪乿丄GR/F傪乽岝棙梡岠棪乿偲屇傫偱偄傑偡丅偙偺夝愅偱傕慜幰偼宍懺揑摿惈丄屻幰偑惗棟揑摿惈傪昞偟偰偄傑偡丅偙傟偼NAR亊LAR偺惉挿夝愅偲偳偙偑堘偆偺偐丄偲偄偆偲丄偙偙偱埖偭偰偄傞偺偼孮棊撪偺條乆側岝娐嫬偵偄傞屄懱偱偡丅摨偠梩柺愊傪帩偭偰偄偰傕丄忋憌偵払偡傞屄懱偲壓憌偵偄傞屄懱偱偼庴岝検偑堘偄偡偓丄LAR偺懡壡偑屄懱偺忬嫷傪椙偔昞偡偲偼尵偊側偄偐傜偱偡丅側偍丄偙偺夝愅偱偼AGR側偳抧忋晹偺惉挿懍搙傪埖偭偰偄傑偡偑丄偙傟偼扨偵孮棊撪偺屄懱偺抧壓晹廳検偼悇掕偟偵偔偄偨傔柍帇偟偰偄傞偩偗偺偙偲偱偡丅
|
棯岅 |
惓幃丠柤徧 |
撪梕 |
傛偔梡偄傜傟傞扨埵 |
摨堄岅 |
|
GR |
growth rate |
扨埵帪娫偁偨傝偺屄懱廳偺憹壛検 |
g d-1 |
|
|
RGR |
relative growth rate |
屄懱廳偁偨傝偺惉挿懍搙 |
g g-1 d-1 |
specific growth rate |
|
NAR |
net assimilation rate |
梩柺愊偁偨傝偺惉挿懍搙 |
g m-2 d-1 丂 |
|
|
LAR |
leaf area ratio |
梩柺愊/屄懱廳斾 |
m2 g-1 丂 |
|
|
LMR |
leaf mass ratio |
梩廳/屄懱廳斾 |
g g-1 丂 |
leaf weight ratio |
|
SLA |
specific leaf area |
梩柺愊/梩廳斾 |
m2 g-1 丂 |
|
|
LMA |
leaf mass per area |
梩柺愊/梩廳斾 |
g m-2 丂 |
specific leaf weight, specific leaf mass |
|
NP |
nitrogen productivity |
屄懱拏慺偁偨傝偺惉挿懍搙 |
g g-1 d-1 丂 |
|
|
PNC |
plant nitrogen concentration |
屄懱拏慺擹搙 |
g g-1 丂 |
|
|
SAR |
Specific absorption rate |
崻廳偁偨傝偺拏慺媧廂懍搙 |
g g-1 d-1 丂 |
|
|
SRL |
specific root length |
崻廳偁偨傝偺崻挿 |
m g-1 丂 |
|
|
|
|
|
|
|
|
偮偯偔乮偨傇傫乯 丂 |
|
|
|
|