光合成の生理生態学講座
はじめに(200004 29)
構成・維持コストを調べることの意味(200004 29)
構成呼吸・維持呼吸(200004 29)
構成呼吸・維持呼吸の理解の問題点(200005 10)
維持コスト(200005 19、200006 10改変)
1 Pathway analysis (200005 26)
2 還元状態からの推定 (200005 31)
3 Heat of Combution (200006 10)
器官間の違い (200006 23)
種間の違い (200006 23・200611 11追加)
生育条件の影響 (200006 23)
植物は光合成により糖を合成し、その合成された糖を使って自らの体を形づくります。とはいっても植物体は糖だけで構成されているわけではなく、糖から様々な物質を合成し、形づくっているわけです。糖から糖以外の物質を合成するためには多くの場合エネルギーが必要です。したがって、植物体形成のためには、材料としての糖と合成のためのエネルギーが必要なわけです。一般に合成のためのエネルギーを構成コスト(construction cost)とよびます。また、エネルギーは合成のためだけではなく、いったん作った植物体を維持するためにも必要です。これを維持コスト(maintenance cost)とよびます。この項ではこの二つのコストについて書きます。
まずは、植物体の構成・維持にはどれだけのコストがかかっているのかは自然な興味の一つと言えると思います。我々は植物の成長速度に着目することが多いのですが、同じ1g増えるのでもコストがかかる植物体ではエネルギーが余計にかかることになります。光合成量が同じだとしても成長速度は違うはずです。
他には、個体間・種間で何らかの「戦略」が異なる可能性がありますが、それにコストが関係している可能性が高いことが挙げられます。80年代から植物体のコストが着目されている理由の一つに、「植物は動物から食べられるのを防ぐために様々な物質を貯め込んでいる」ということがあります。前述のように、コストがかかる植物体を作ることは成長速度を相対的に遅くすることになるわけですが、そういう不利益を払っても、食べられない、という利益を得られるのであればかえって有利かもしれません。ある「戦略」を採るときには、常にその利益と不利益がつきまといます。構成・維持コストを調べることは、その「戦略」の不利益を明らかにすることに役立つことが多いと期待できます。
この業界にいる人なら構成呼吸・維持呼吸という言葉を知っている人は多いと思います。ここでは歴史から振り返ることにしましょう。
概念的には、構成呼吸・維持呼吸というのは以下のようにとらえられています:呼吸で生産されたエネルギーによって払われるという観点に立ち、それぞれのコストを呼吸量に換算したものが構成呼吸・維持呼吸。
実験事実からこの考え方を最初に提唱したのは、戦後東大植物学教室にいた田宮さんだという話を聞いたことがあります。ゾウリムシの呼吸速度を調べた結果と聞いていますが、残念ながら私は文献までは知りませんし、実際現在この分野では引用されているのを見たこともありません。 実質上最初の発見者とされているのはMcCree(1970)です。McCree(1970)はシロツメクサの光合成と呼吸を個体丸ごと測定する、ということをしました。ここで、彼は面白い測定の仕方をしました。まず、一日めは100 W m-2 の光をあて、昼中光合成を測り続け、日光合成量を測定します。その日の夜には光を消し、夜間の総呼吸量を測定します。次の日は、同一個体について同様の測定を光強度を変えて測定しました。それを何度か行い、彼は様々な光強度での光合成量と個体重と呼吸量の間には相関があることに気が付きます。実験結果をまとめると、以下の式が得られました。
個体呼吸量 = k 光合成量 + c 個体重
ここで、kとcは係数です。ここで、k×光合成量に相当する呼吸量を「構成呼吸」、c×個体重に相当する呼吸量を「維持呼吸」と呼ぶことになります。この式は後に両辺を個体重で割り、光合成量のかわりに成長速度を用いて、以下の式で使われるようになりました。
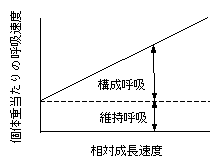
r = k RGR + c
ここで、rは個体重当たりの呼吸速度、RGRは相対成長速度(個体重あたりの成長速度)です。kを構成呼吸係数、cを維持呼吸速度(維持呼吸係数)と呼びます。この式は以下のような図になります。
概念的には以下のように説明されます。成長、すなわち新たな物質の構成には生成エネルギーがかかり、呼吸速度には構成量すなわち成長速度が高いほど高くなる成分(k×RGR)があり、これを構成呼吸と呼びます。一方、すでに作ってしまった物質の維持にもエネルギーは必要なはずです。その分の呼吸は構成呼吸を除いた部分の呼吸、つまり成長速度0のときの呼吸速度として認識できるはずです。ということで、Y切片cを維持呼吸速度とみなすわけです。
ちなみに、構成呼吸・維持呼吸については1970年代から都立大でも盛んに研究が行われていました(Yokoi et al. 1978, Kimura et al. 1978, Mariko 1988)。
呼吸速度が上に示した式で表される、ということそのものは、経験的かつ普遍的な知見なので、それはそれでいいのですが、それで本当に構成と維持のコストを理解した気になっていいのか?という問題は残ります。実際に、いくつか問題点を指摘することができます。
いずれどこかで詳しく書くと思いますが、多くの植物では細胞中の糖類の濃度と呼吸速度の間に相関があることが示されています(Azcon-Bieto and Osmond 1983)。呼吸系も酵素反応ですので、基質である糖の濃度に呼吸速度が依存することは何の不思議もありません。さらに、光合成をたくさんした葉ほど細胞内糖類含量が高いことも当然です。したがって、McCreeの結果が意味するのは単に光合成量が多い個体では糖濃度が高いために呼吸速度が高いのであって、構成コストとはまた別の問題であった可能性はあります。この点をちゃんと区別した研究があるのかどうか私は知りません。
呼吸において最も多くのATPを生産するのはミトコンドリアの電子伝達系です。植物(と昆虫の一部)には二つの電子伝達系があることが知られています。一つはATP生産のための電子伝達系で、cytochrome pathway と呼ばれます。文字どおりもう一つの電子伝達系がalternative pathway と呼ばれる系です。cytochrome pathway はシアンで阻害することができるのですが、alternative pathway はシアンで阻害されないので、シアン耐性呼吸と呼ばれたりもします。この系がalternativeと呼ばれる理由は、この系に入った電子はATP生産に貢献しないためです。Azcon-Bieto et al. (1983) の研究では、糖分濃度上昇による呼吸速度の上昇のかなりの部分がalternative pathway 由来であることを示しています。したがって、呼吸速度が上がったからといってそれが必ずしもATP上昇やコスト上昇と関係がない可能性があります。
alternative pathway と維持呼吸・構成呼吸の関係については H Lambers のグループによって、80年代に精力的に調べられました。Lambersが着目したのは主に根で、維持呼吸・構成呼吸の他に、イオン吸収に必要なエネルギーコストも考慮したエネルギー分配について調べています。集大成とも言うべき?論文を一本だけ紹介しておきます(van der Werf et al. 1988)。これについては機会があれば書きます。
細胞はその体を維持するために常にエネルギーを必要とします。でも、具体的には何に必要なのでしょうか? 維持コストに関わる研究のほとんどは上記の維持呼吸関連のもので、呼吸によるエネルギー消費量を求めたにすぎません。この問題をマジメに考えたのは、私の知る限りPenning de Vries (1975) が最初です。彼は細胞の維持に必要なのは、1)細胞構成物質のターンオーバー、2)膜間イオン勾配の維持、3)来る環境変化へ適応可能なように細胞をactiveに保っておくこと、と考えました。
1)は細胞内におけるある物質の分解と再合成を考えています。タンパク質のターンオーバーが中心のようです。といっても彼自身がタンパク質のターンオーバーを測定したわけではなく、文献値から推定したわけです。結果は、バイオマス1gを維持するのに、一日あたり7-13mgのグルコースを必要とするとしています。
次に2)ですが、生体膜は膜内外で物質を分けておくという役割を担っています。特にイオンは特定のトランスポーターを介さない限り膜を透過できないとされていますが、実際には結構ぽろぽろイオンを透過してしまうようです。放っておいては役割を成さないので、エネルギーを使って漏れ出たイオンを回収する、ということをする必要があります。例えばナトリウムイオンをひとつ細胞外にかき出すためにはATPがひとつ必要です。主要なイオンについて膜から漏れ出す速度とそのイオンを元に戻すためのエネルギーを文献値から計算することができます。その結果は、バイオマス1gを維持するのに、一日あたり6-10mgのグルコースを必要とするとしています。3)は種によって違うと考えられますので、とりあえず1)と2)を近本的な維持コストとして考えることができます。
Penning de Vries (1975) は、これによって求めた維持コストと他研究が測定した維持呼吸を比較する、ということをしました(この論文にはいろいろな研究の維持呼吸速度の表が載ってます)。それによると、細胞はバイオマス1gを維持するのに、一日あたり8-60mgのグルコースを必要とするようです。Penning de Vriesの計算よりはちょっと多いようです。彼は生育条件の問題などがある(生育条件によってタンパク質含量などが違うので維持コストが異なると予測される)ためかもしれない、と書いてますが、まあ未だにわかっていないことは確かです。
この問題の研究はこの後ほとんど進んでないようです(Amthor (1994) も何も書いてないなあ)。20年以上の時を経て?大阪大の野口さんがこの分野に手をつけ始めたようでいるようで、「葉っぱの呼吸のうちこの分はタンパク質のターンオーバーで消費されてます」という図が野口(1999)に載っています。
構成コストを測定するための手法はいくつかあります。
これは、材料とする植物体にどういう物質がどれだけあるかを測定し、それぞれの物質の生成コストを考慮することにより植物体の生成コストを計算するものです。Chapin (1989) の方法を説明しましょう。彼は植物体はおおざっぱにわけて、タンパク質、リグニン、タンニン、脂質、炭水化物、繊維からなると考えました。そして様々な生育型(草本、コケ、木本など)の葉と茎におけるこれらの物質の割合を測定しました。彼によれば、この6種類の物質で植物体の95%を説明できたということです。次に、それぞれの物質の生成コストを計算します。これは、代謝経路から推定しています。各代謝ステップでどれだけエネルギーが必要かはほとんどの代謝経路でわかっていますから、例えば1gのリグニンを作るために、必要なATP、そしてそのATPを呼吸で生産するために必要なグルコースの量を推定することができます。炭水化物や繊維はコストがかからない物質で、1g作るのに必要なグルコース(炭素骨格+生成エネルギー)は1.1-1.2g程度です。しかし、タンパク質やタンニンはその約1.5倍、リグニンは約2倍、脂質は約3倍コストがかかります。これらの各物質の量とコストから植物体のコストが計算できます。Chapin (1989) によると、彼が調べた範囲(アラスカの様々な生育型の植物)では、どの植物でも葉・茎ともだいたい1gあたり1.5g前後で、特に種間差に傾向はなかったそうです。
問題点は、まず各物質の測定に手間がかかる上に正確に測定することが難しいことでしょう。そんなわけで過去にはあまりこの方法でコストを推定した仕事は多くなかったのですが、最近H PoorterやVillarのグループからの報告が増えているようです(Poorter et al. 2006, Villar et al. 2006)。
植物体の元素組成から構成コストを推定するものです。ある物質を作るときに必要な炭素と電子を供給するために必要なグルコースの量を計算します。McDermitt and Loomis (1981) により考案されました。
ある物質CcHhOxNnSsを考えると、reduction level: r = 2x - h - kn -ms という式を導くことができます。kとmは、それぞれNとSの由来の気質の酸化数(oxidation number)です。窒素の場合は、根から吸収された窒素が硝酸イオンの場合は5で、アンモニウムイオンの場合は-3です。硫黄の場合は、硫酸イオンだと6で、硫化水素だと-2です。このrが0以下であるということは、もとの状態(吸収された状態)からこの物質を作るために還元力(電子)を必要とした、ということです。逆に0より大きい場合は、物質生成の間に電子を放出したということです。ちなみに、グルコースはr=0です。cだけ炭素を持っている物質を1mol作るためのコストは以下のように定義できます。炭素骨格として、グルコースの炭素(1分子あたり6つ)を使うので、c/6molのグルコースがまず必要です。さらに、還元力が必要になります。1molの電子を放出するために1/24molのグルコースが必要だと仮定すると、構成コストは
グルコース当量(glucose equivalent)= c/6 - r/24
となります。
Mcdermitt and Loomis (1981) の論文はよく引用されるのですが、この方法を使ってコストを推定した論文は見たことないですね。しかし、彼らが考案したglucose equivalent という概念は以後この関係の仕事で標準的に使われることになります。上のChapin (1989) の仕事もそうです。
ある物質を燃焼したときに出る熱量をthermometerによって測定し、この値がglucose equivalentと相関があることを利用します。簡単に言うと、物質のカロリーを測っているわけです(いわゆるカロリーが高い食品、というのはエネルギーコストが高い)。Williams et al. (1987) によって確立されました。彼らの論文によれば、glucose equivalent (GE、g glucose/g dry mass) とheat of combution (dHc、kJ/g dry mass) の間には以下の関係があります。
GE = (0.06968 dHc - 0.065)(1-A) + kN/14 × 180.15/24
ここで、Aは灰(燃えカス)の重さ(g/g dry mass)、kは窒素源のoxidation state(g N/g dry mass)、Nは窒素含量(g N/g dry mass)、14は窒素の原子量、180.15はグルコースの分子量、24はグルコース1モルから出てくる電子の数です。
この方法の利点は、なんといっても簡単に測定できることです。1の方法にしても2の方法にしてもめんどうな生化学手法によって構成物質の組成を明らかにしなくてはいけません。その点この方法は物質を一度燃やしてしまえばいいだけなわけですから。近年の仕事ではほとんどの研究がこの方法で構成コストを測定しています。
90年代から、構成コストの研究は増えています。器官あるいは種間でどのような違いがあるかは興味深いところです。これまでに、Griffin (1994)、Poorter (1994)、Poorter and Villar (1997) が文献サーベイを行っています。ここでは、最も新しいPoorter and Villar (1997) を少し紹介しましょう。
様々な文献データを平均してみると、栄養成長器官(つまり、葉・茎・根)では、葉がやや高い傾向にあるようです(平均GE=1.5)。茎は1.45、根は1.33と低くなっていますが、根の測定例は少なく、まだ不確定なようです。
リグニンを多く含む木本の茎は構成コストが高いような印象を受けますが、平均すると1.5とそれほど高いわけではありません。これは、コストが低いセルロースの類も含まれているためです。
繁殖器官では、種子のコストが種によって大きく異なり、高い種ではGEが2を超えます。これは脂肪・タンパク質・デンプンと種子が貯蔵する物質が種によって異なることを反映しています。が、平均すると1.53と葉とそれほど変わらない値になっています。果実の類は平均が1.41で、種子に比べればコストが低いものが多いようです。
Poorter (1994) は文献サーベイから種間では系統だった構成コストの違いはない、と結論していました。その後文献が増え、再検討した結果、今度は前の結論を覆したようです(Poorter and Villar 1997)。葉の構成コストGEは、草本植物157種で1.45、落葉樹36種で1.52、常緑樹35種で1.58だったそうで、これは有意差があるのだそうです。この結果はなかなか興味深い結果です。といいますのは、Chapin (1989) やWilliams et al. (1989) では、葉の寿命が長い植物では葉のコストが高いのではないか、という仮説があったのです。葉の寿命を長くするためには、食植者や物理的撹乱に対する防御のために葉を丈夫にしたり二次代謝物質を貯め込んだりと、コストを高くする必要があるのではないか、と考えられていたのです。しかし、彼らの調査ではそのような結果は出ていませんでした。Poorter and Villar (1997) の結果は、葉の寿命が長い植物ほど構成コストが高いことを示唆し、ChapinやWilliamsらの仮説がある程度正しかったことを示しています。しかし、彼らの実験ではそのような結果が得られなかったこと、1994年当時の調査では差が出ず、1997年の調査では差が出た、などということは、差はあってもそれほど大きいわけではない、ということを意味するのだと思います。
Villarらは2006年にさらにつっこんだ解析の結果を発表しました。ここでは文献サーベイではなく自分たちで測定をしています。特に着目に値する点は、一つの単系統から常緑と落葉を二種選び、この組み合わせを8つの組み合わせ(単系統)について調べることにより系統の影響と落葉・常緑の違いを分離して調べたところです。また、化合物を同定して炭素コストをpathway analysisによって推定していること、も先行研究と異なる点です。その結果は、「落葉常緑の差は小さく、コストや化合物組成の違いは系統間の違いのほうが大きい」というものだったようです。
以下に記すのは、同一種の葉の構成コストが生育環境によってどう変化するかです。
高CO2で育てると、多くの場合コストは下がるようです。これは、高CO2生育の植物は炭水化物の蓄積が多く、タンパク質含量が少ないことによるようです。
光環境の影響は研究によって様々なようですが、強光条件でコストが上がることが多いようです。
栄養条件では、富栄養条件ほどコストが上がるようです。これは、タンパク質含量が多いことで説明できるようです。